Background: Retinopathy of prematurity (ROP) is considered as the most common reason for blindness in children, particularly in preterm infants. The disease is characterized by the dysregulation of angiogenic mechanisms due to preterm birth, leading ultimately to vascular abnormalities and pathological neovascularization (NV). Retinal detachment and vision loss could represent a concrete risk connected to the most severe forms of ROP, also characterized by inflammation and retinal cell death.
Methods: During the last decades, many animal models of oxygen-induced retinopathy (OIR) have been recognized as useful tools to study the mechanisms of disease, since they reproduce the hallmarks typical of human ROP. Indeed, modulation of retinal vascular development by exposure to different oxygen protocols is possible in these animals, reproducing the main pathological phenotypes of the disease. The easy quantification of abnormal NV and the possibility to perform electrophysiologic, histological and molecular analyses on these models, make OIR animals a fundamental instrument in studying the pathophysiology of ROP and the effects of novel treatments against the disease.
Discussion: Here, the most commonly used OIR protocols in rodents, such as mice and rats, are described as well as the main pathological outcomes typical of these models. Despite their limitations and variables which should be considered whilst using these models, OIR models display several characteristics which have also been confirmed in human patients, validating the usefulness of such animals in the pre-clinical research of ROP.
Retinopathy of prematurity (ROP) is the most common cause of blindness affecting preterm infants (1). Similarly to other proliferative retinopathies, ROP is characterized by the disruption of normal angiogenic mechanisms leading to retinal neovascularization (NV) (1). In premature infants, the interrupted development of the retinal vasculature at the time of birth, together with an initial hyperoxia-induced microvascular obliteration, determine the onset of a peripheral avascular area (2). These vascular changes result in an increased hypoxia in the avascular areas, due to the gap between oxygen supply and increased metabolic demands of the developing retina (2). As a result, the massive upregulation of pro-angiogenic factors promotes the abnormal retinal NV, characterized by the formation of leaky, non-organized vessels, called pre-retinal tufts, which grow into the vitreous cavity and lead to visual loss following retinal detachment (2). Pre-retinal tufts can regress in case of adequate oxygen administration (2). However, although about 90% of ROP infants suffer from only mild visual impairment, patients with untreated NV are still at an elevated risk for progression to more severe conditions, promoting impaired vision and blindness (1). Several multifactorial agents characterize the pathogenesis of ROP, involving the crosstalk among different pathophysiological factors, such as hypoxia, inflammation and oxidative pathways (3).
Animal models of oxygen-induced retinopathy (OIR) are popular models widely used to reproduce ROP and investigate pathologic NV in the retina, as well as the involved molecular mechanisms and new putative therapies. Several common OIR models were established in different species, such as feline (4), murine (5,6), and canine (7), all of which mimicking ROP in humans. A fundamental prerequisite for an animal OIR is an immature retinal vasculature at birth with an active physiological angiogenesis, similar to the human retina before 31 weeks of gestation (1). The exposure to constant or cycling hyperoxia in OIR models leads to the arrest/retardation of normal retinal vascular development, vascular dropout and following retinal NV, thus reliably reproducing the two phases of ROP (1). Although the reproducibility of NV is different across species, the easy induction of NV, together with the rapid visualization and quantification of pre-retinal tufts, makes OIR models useful tools for the study of mechanisms and potential therapeutics against ischemic retinopathies, including ROP (8). To date, the most commonly used OIR models are performed in rodents, such as mice and rats, which are discussed in the following sections. Smith’s mice OIR (6) and Penn’s rat OIR model (5) protocols are described. Following the description of the protocols, a brief overview of several outcomes deriving from the application of the procedures described below is given. In this respect, practical suggestions about techniques which could be used for each aspect are provided, as well as a brief description of exemplificative results deriving from the application of such methods.
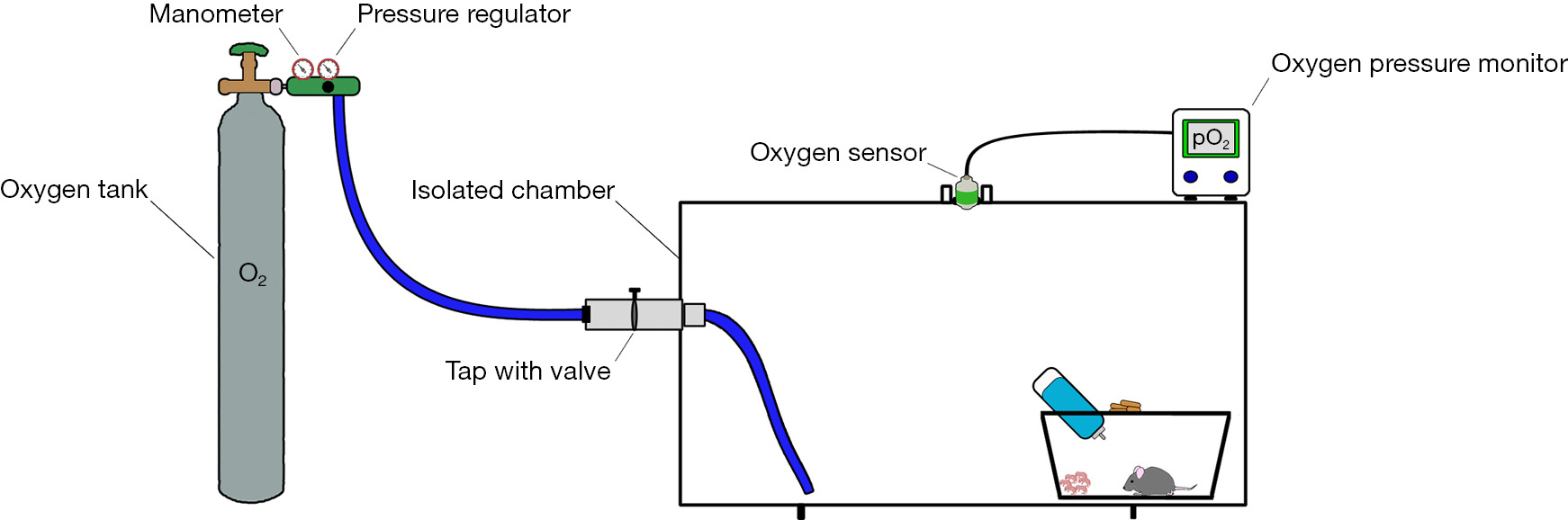
A schematic representation of an example of how a custom-made chamber could be set up is shown in Figure 1.
A schematic representation of the protocol is shown in Figure 2A.
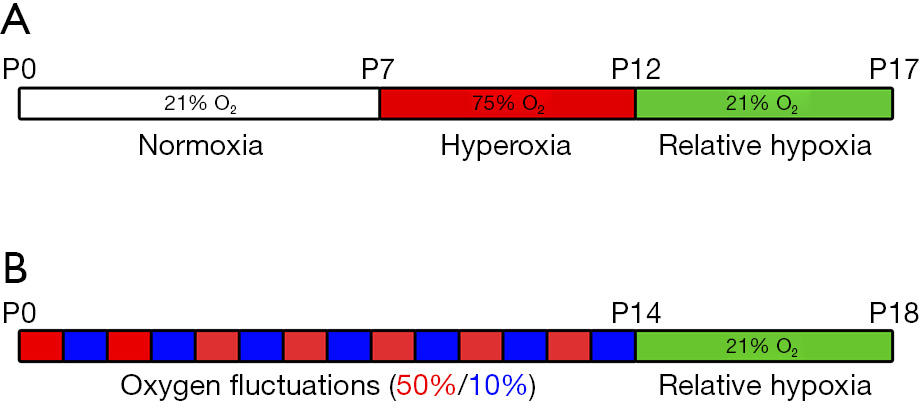
A schematic representation of the protocol is shown in Figure 2B. The protocol described here is the result of several changes applied over time to have a model mimicking more closely the clinical settings of ROP. From an initial exposure of the rat pups at continuous 80% O2, the hyperoxia exposure paradigm was changed to 80%/40% cycles and, finally, to alternating 50%/10% before keeping the animals to room air (13). Moreover, also the timing of the oxygen exposures has been progressively changed over the different protocols (13). The 50/10 rat model described here has been considered to more closely reflect the arterial blood gases experienced by a sick premature infant and result in a greater retardation of retinal blood vessel development, as well as in an increased NV severity compared to other rat models (13).
ROP is known to result in long-term effects on vision, which have been observed in patients even after the active phases of the disease (17). Functional alterations of the retina following an OIR protocol could be assessed by electroretinogram (ERG). In particular, a full-field ERG could be used to evaluate the activity of the entire retina in animals after the exposure to OIR protocols (18). This non-invasive analysis can be conducted in vivo under physiological or nearly physiological conditions through electrodes collocated on the corneal surface. The positive and negative components of the recorded ERG wave indicate the sum of the potentials originating from different stages of retinal processing after a provided light stimulus (18). ERG analyses should be performed under systemic anesthesia, and the further application of local anesthetics is recommended to avoid blinking. ERG in developing pups should be recorded at later stages than P14, given that the animals open their eyes by the end of the second post-natal week. Thus, it is recommended to perform functional analysis at least P17/P18, when an adequate eye opening has occurred favoring the correct electrodes positioning and the wide opening of the vision field. To evaluate retinal function, a dark-adapted scotopic ERG could be recorded (18). A scotopic ERG wave usually comprises of two components, the initial negative-going a-wave (mainly from photoreceptors) and the following positive-going b-wave, generated by post-receptor cells (18). Evidence of impaired ERG responses has been reported both in OIR mice and rats. In particular, functional impairment can be observed during the hypoxia phase, evidenced by the reduced a- and b-wave amplitudes in OIR animals (19,20). Moreover, functional deficits in OIR models persist even after vascular regression (21,22).
The visualization of retinal vasculature on flat mounts represents the gold standard method to evaluate any sign of vessel alteration induced by the OIR protocol. Retinal samples could be fixed and labelled with specific molecules to recognize endothelial markers, thus ultimately providing a tool to label retinal vessels (Figure 3). The most commonly used vascular labelling is performed with isolectin B4 (IB4), which allows vascular visualization by targeting the surface of endothelial cells (14). The whole flat mount analysis allows the experimenter to have a full contemporary view of the retinal vascularization, thus providing an ideal instrument to quantify OIR vascular outcomes (11). Indeed, avascular areas and pre-retinal tufts could be quantified through image-processing software (i.e., Photoshop, Image J) starting from a retinal flat mount image acquired through the microscope. For example, vaso-obliterated areas could be measured by outlining the avascular area related to total retinal area (23,24). On the other hand, pre-retinal tufts could be measured either manually or semi automatedly and be expressed as a percentage of total retinal area (23). Alternatively, intravitreous NV in the rat OIR model could be analyzed by counting the number of clock hours involved or summing areas of NV (24).
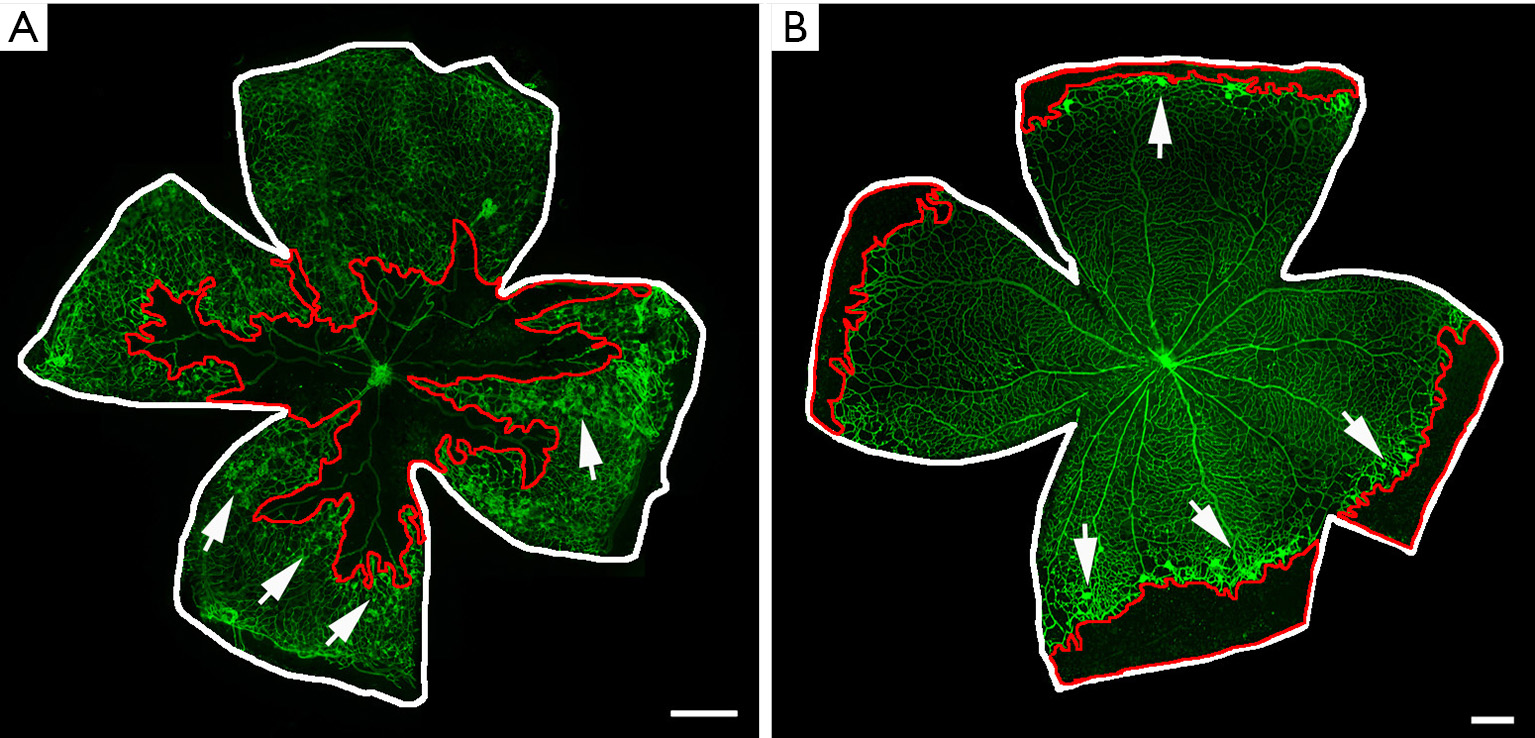
In an OIR mouse model, a central avascular area can be observed at P12, as a consequence of the hyperoxia-mediated vaso-obliteration (11). As a response to ischemic conditions after returning to room air, the significant NV leads to the growth of neovascular tufts from P14 to onward, reaching the peak intensity at P17 (11). Thus, the visualization and quantification of NV on retinal flat mounts is mostly performed at P17. After this time point, abnormal NV begins a spontaneous gradual regression, completely disappearing by approximately P25, when almost neither vaso-obliterated areas nor NV are visible (23).
Unlike the mouse model, in the rat OIR retinas the exposure to cycling levels of oxygen slows the retinal vascular development in peripheric areas. Indeed, by P14 the 50/10 OIR retinas display a peripheral avascular area, compared to the fully vascularized retinas of room air pups (24). This time point, which correspond to the end of oxygen fluctuations, could be used to analyze the formation of the peripheral avascular area, a typical hallmark of the protocol success. In the rat OIR model, NV occurring at the boundary between vascularized and avascular areas in the midperipheral retina reaches its peak at P18/P20 (24). Similar to the mouse model, NV recovers by P25, thus to observe differences in retinal vascularization it is recommended to end the experiment at P20 (14).
Vascular dysfunctions characterizing OIR retinas occur as a consequence of the dysregulated expression of pro-angiogenic factors due to hypoxia conditions (2). Molecular analyses represent a useful tool to assess the changes in the levels of pro-angiogenic factors occurring at either transcriptional or translational level during the OIR protocol. It should be considered that the small dimension of neonatal pup retinas may hinder the molecular experiments, due to the unavailability of sufficient tissue to provide acceptable material for the analysis. Thus, samples containing pooled retinas from more than one animal are often required to obtain adequate protein or RNA levels for quantification (11).
In a mouse OIR model, a downregulation of pro-angiogenic factors, such as vascular endothelial growth factor (VEGF) and erythropoietin (Epo), is observed at P12 after hyperoxia (1,23). In contrast, during NV an increase in expression of pro-angiogenic factors, such as VEGF, Epo, insulin growth factor 1 (IGF-1), fibroblast growth factor 2 (FGF2), heparin-binding EGF-like growth factor (HB-EGF), and hepatocyte growth factor (HGF), as well as the activation of VEGF, Epo and IGF-1 receptor pathways, are reported as a consequence of hypoxia (23,25-29). In contrast, in the rat OIR model an upregulation of VEGF and VEGFR-2 levels has been reported both at the end of oxygen fluctuations (P14) and retinal NV (P18) (24,30).
The activation of inflammatory mechanisms is another pathological feature of OIR models. Indeed, increasing evidence shows that a local and/or systemic inflammation might contribute as a risk factor in ROP, and the increment of inflammatory molecules has been demonstrated to perturb retinal vessel development and induce pathological features of ROP (31). The increased expression of pro-inflammatory molecules could be assessed through molecular analyses from retinal extracts, both at transcriptional and protein levels. In this respect, in an OIR mouse model several pro-inflammatory molecules have been observed to increase during the early stages of hypoxia, with a following decline during the vascular regression phase (32).
Furthermore, inflammation is often accompanied by Müller cell gliosis and microglial/macrophages activation, increasing the wide spectrum of mechanisms damaging OIR retinas (33). Indeed, these cellular types have been demonstrated to be involved in the production of inflammatory molecules and the mediation of retinal damage (34). An immunohistochemical analysis on retinal sections could be used as an example to visualize these cell types in the retina during its inflamed status. In this respect, markers labeling glial cells could be used to visualize the localization of the cells and their activation state. For example, the frequently used marker to label gliotic Müller cells is glial fibrillary acidic protein (GFAP), whereas ionized calcium-binding adapter molecule 1 (Iba1) is the most commonly used marker to detect retinal microglial cells and recruited macrophages. The upregulation of GFAP, as well as the increase in GFAP positive processes among the entire retina, are documented in an OIR model during hypoxia, suggesting the activation of Müller cells (35). Moreover, an increase in GFAP immunoreactive processes underlining the activation of Müller cells is documented also in the OIR rat model (36). Consistently, an increase in ameboid shaped Iba1 positive cells, resembling the morphology of activated microglia/macrophages, has been described in mice OIR retinas (35). In this respect, when performing microglial/macrophagic labeling, it is recommended to add the use of other markers which could indicate the subpopulation and the activation state of microglia/macrophages, since different phenotypes of these cells might help either retinal NV or revascularization and disease resolution (32).
Vascular alterations and retinal inflammation could lead to retinal cell damage, ultimately resulting in retinal cell death. The overall increase in retinal cell death during an OIR protocol could be indicated by the reduction in the thickness of retinal layers, assessed through histological analysis after the labeling of cell nuclei by 4',6-diamidino-2-phenylindole (DAPI) counterstaining or through histological stains to visualize retinal layers. In addition, to assess morphological alterations on photoreceptor outer segments, histological analyses using specific markers labeling such portions could be useful to evaluate putative shortening or disorganizations of these segments (35). A decrease in total retinal thickness is observed in both mouse and rat OIR models, especially in the inner retinal layers which seems to be the most affected by the changes in oxygen pressures (19,37). Nonetheless, some evidence reported also damage to the outer retina, with reduced outer nuclear layer (ONL) thickness and decreased photoreceptor density, especially in the rat model from later stages (P60) (38,39). Moreover, a shorter and disorganized structure of photoreceptor outer segments has been reported in both models, which might partially explain the functional impairment of the ERG a-wave (35,40). Furthermore, evidence of apoptotic microvascular cells, such as endothelial cells and pericytes, has been documented (41). The alterations in retinal thickness persist also during late stages after oxygen exposure (21).
Emerging evidence has underlined that the choroid is also extensively affected during the development of ROP both in animal models and in humans (33). The effects derived from the application of OIR protocol on choroidal vasculature could be evaluated by histological analyses to assess choroidal thickness. An example of such analysis could be performed by the staining of retinal/choroidal sections through markers labeling the endothelial cells of choroidal vessels (38,39). Alternatively, molecular analyses of cell death markers, at either transcriptional or protein levels, could be performed from choroidal tissues extracts to assess some basic molecular events associated with choroidal involution (38).
In the rat OIR model, a choroidal thinning was observed at P14 following oxygen fluctuations, which persisted during retinal NV phase and over time up to P210 (38,39). Concomitantly, the upregulation of cell death markers has been observed during oxygen fluctuations and NV phases (38).
ROP is one of the most common reasons of blindness occurring during prenatal stages (3). Therefore, an appropriate model able to reproduce faithfully the main features of the disease, such as vascular dysfunction, inflammation, retinal damage, choroidal involution and functional impairment, might represent a useful tool to study pathophysiological mechanisms and possible therapeutical strategies. In this context, the OIR models provide a bona fide representation of the most relevant events associated with ROP, thus offering a valid instrument to analyze the mechanisms involved in the disease and some hypothetical resolutions. Actually, most of the information about molecular mechanisms of the disease, as well as the effects of several drugs, come from these models, underlining the importance of these animals in ROP research.
The most important prerequisite of an animal OIR is represented by an immature retinal vasculature at birth, whose postnatal development could be easily modulated to mimic vascular abnormalities typical of human ROP (1). Unlike humans, where the retinal vasculature matures before a baby is born term, these animals develop their retinal vascular system during the first postnatal stages, thus resembling the condition of a human preterm retina (1). In this respect, the exposure to either constant hyperoxia or oxygen fluctuations during these early phases of development in different species reproduce an initial vaso-obliteration, followed by abnormal NV during the return in normoxic atmosphere, reliably reproducing the phases of human disease (1). Since the neovascular response is very reproducible and quantifiable, the study of the effects of antiproliferative and antiangiogenic treatments could be easily quantified in these models. Considering that OIR models are not only characterized by vascular alterations typical of the human disease, but are also affected by inflammation, retinal cell death, choroidal degeneration and functional loss, these models might provide all the elements which could be considered as hallmarks to evaluate the effect of a putative protective treatment. Indeed, these emerging pathophysiological events have been also observed in ROP patients, where local and systemic inflammation, choroidal thinning and long-term functional impairment might occur in the pathogenesis of the disease (17,33). Therefore, OIR models might represent a helpful tool to study these mechanisms and the efficacy of novel promising therapeutical strategies applied to these factors more deeply.
The OIR mouse model by Smith (6) and the rat model by Penn (5) are the most extensively used for studying the mechanisms involved in pathological angiogenesis, as well as the effects of various angiostatics or neuroprotectants (8). However, although both models could reproduce basic characteristics of the disease, there are some differences in phenotypic outcomes, especially in vascular defects. Indeed, while in the mouse OIR model the vaso-obliteration during the exposure to hyperoxia involves central vessels, the oxygen fluctuations lead to the formation of an avascular area in retinal periphery (8). Therefore, in this aspect, the rat OIR model reproduces more faithfully what happens in human ROP, providing a more reliable model to use when evaluating preventive effects aiming at promoting the revascularization of non-perfused periphery. Nevertheless, the mouse OIR model reliably reproduces the pathologic events occurring during ischemia-induced NV (8). The levels of observed NV are higher compared with the rat model, supporting its use in the screening of antiangiogenic compounds (8). Moreover, another advantage of using mice is the relatively higher ease of genetic manipulation to study directly the role of different genes in NV. For instance, mice OIR models carrying a deletion of beta-adrenergic receptors 1 and 2, were useful to study the role of adrenergic system in pathological angiogenesis and find a promising therapeutical target of disease (42).
Despite the wide usage of these models in pre-clinical research of ROP, there are several limitations connected to important factors which could influence the variability of OIR outcomes. Indeed, strain-dependent differences in terms of NV severity have been reported both in mice and rats, where the extension of avascular areas and NV differs among strains (11,43). Moreover, variability is further promoted if animals belonging to the same strain come from different vendors (11,13). In addition, another source of variability can be found among different mouse litters, since different numerosity and nursing conditions could lead to diverse health of pups, postnatal weight gain and then OIR phenotype severity (11,13). Therefore, all these variables should be addressed in order to obtain a higher reproducibility in obtaining OIR phenotypes. As another relevant point, other complications of premature birth are not reproduced by OIR models since protocols are conducted postnatally with healthy animals without all the comorbidities typically of human preterm infants (3,11). In this respect, healthy animals display a mature lung physiology, which might derive in systemic (and presumably retinal) oxygen tensions that might not be similar to the clinical setting. The OIR rat model has addressed this issue by using fluctuating oxygen exposures in ranges that produce more clinically representative arterial oxygen pressures. However, the timing is much different and less random from that connected with the variable respiratory function in preterm infants with immature lungs (44). Finally, another limitation of the most commonly used OIR models in rodents is represented by the fact that these animals are not characterized by some features of ROP, such as retinal detachment (8).
Despite OIR models are characterized by several factors which could influence the variability in their outcomes, the appliance of OIR models in ROP research has played a pivotal role so far, and with the new advances in genetic manipulation, a deeper understanding of the role of new genes in ROP pathogenesis could be possible. Moreover, the versatility of these models due to the reproduction of a reliable human-like NV phenotype, makes the OIR models useful for the study of other ischemic retinopathies such as diabetic retinopathy (DR). Indeed, the current rodent models of DR (either genetic or drug-induced) are able to reproduce only the early non-proliferative stages, whereas they fail to develop the NV associated to the proliferative phase, likely due to the short life span of the animals (8). Since the mechanisms behind NV and tuft formation are similar between ROP and DR, OIR models could represent a valid alternative to study the dynamics characterizing the proliferative stages of DR and the potential of antiangiogenic drugs appliable to both the diseases. Therefore, the potential of these models to be a compensatory model to study the angiogenic response in other proliferative retinopathies, makes OIR models even more valuable in research of retinal diseases.
In conclusion, OIR models display several characteristics also confirmed in human patients, which make these animals a bona fide tool in the pre-clinical research of ROP and other proliferative retinopathies. Nonetheless, there are many limitations and many variables which should be considered whilst using these models to obtain proper results. In this respect, this study protocol tries to provide starting guidelines critically introducing the basic knowledge of OIR protocols, the related outcomes and its variables and limitations. This study protocol has the aim to provide a critical tool to colleagues who want to start with the OIR studying, and a helpful instrument for a critical beginning of their work, which will be further deepened by the literature regarding the specific topic they will choose.









