Background: Retinal degeneration is a common feature of several retinal diseases, such as retinitis pigmentosa and age-related macular degeneration (AMD). In this respect, experimental models of photo-oxidative damage reproduce faithfully photoreceptor loss and many pathophysiological events involved in the activation of retinal cell degeneration. Therefore, such models represent a useful tool to study the mechanisms related to cell death. Their advantage consists in the possibility of modulating the severity of damage according to the needs of the experimenter. Indeed, bright light exposure could be regulated in both time and intensity to trigger a burst of apoptosis in photoreceptors, allowing the study of degenerative mechanisms in a controlled fashion, compared to the progressive and slower rate of death in other genetic models of photoreceptor degeneration.
Methods: Here, an exemplificative protocol of bright light exposure in albino rat is described, as well as the main outcomes in retinal function, photoreceptor death, oxidative stress, and inflammation, which characterize this model and reproduce the main features of retinal degeneration diseases.
Discussion: Models of photo-oxidative damage represent a useful tool to study the mechanisms responsible for photoreceptor degeneration. In this respect, it is important to adapt the exposure paradigm to the experimental needs, and the wide range of variables and limitations influencing the final outcomes should be considered to achieve proper results.
Trial Registration: None.
Light represents a double-edged sword in the retina, since it is fundamental for visual perception but results in harmful effects on retinal cells under excessive exposure (1). Indeed, intense light exposure to the retina could contribute to the rapid increase in reactive oxygen species (ROS), thus promoting the onset of oxidative stress if ROS overproduction exceeds the endogenous antioxidant capacity (2,3). The ROS-mediated oxidation of cell biochemicals, such as lipids, proteins, and DNA, might result in the alteration of cell viability, further compromised by oxidative stress-induced inflammation, gliosis, and microglial activation (4). These mechanisms ultimately influence the activation of apoptosis in retinal cells, leading further to functional loss (5). In this respect, photoreceptors have been demonstrated to be the first cell type to show the signs of damage associated to prolonged intense light exposures (6). Since photoreceptor degeneration characterizes several retinal diseases, experimental models of photo-oxidative damage, or retinal light damage (LD), have been used to mimic human retinal degenerations arising from environmental and genetic insults, which are based primarily on similar morphological and molecular outcomes. For example, the degeneration of photoreceptors observed in LD models results in the functional loss observed in many retinal disorders, including retinitis pigmentosa and age-related macular degeneration (AMD) (7). Furthermore, oxidative stress and chronic inflammation are also pathophysiological events involved in these diseases (8,9). The advantage of these models in studying photoreceptor degeneration is that light exposure can be regulated according to experimenter necessities, to control the induction, timing, and extent of photoreceptor death. Indeed, the exposure to intense visible light simultaneously triggers cell death mechanisms in a large number of photoreceptors, leading to the synchronization of photoreceptor degeneration facilitating the detection and the analysis (10). Therefore, the use of LD models instead of models of inherited retinal degeneration, where cell death proceeds rather slowly and progressively, could be functional to study minimal changes in levels, localization, modifications and activity of factors involved in the regulation of photoreceptor damage (10). Moreover, the effects of a putative either preventive or curative treatment could be easily assessed in a relatively short time frame, given the large-scale degeneration and the possibility to modulate the severity of damage. In this section, an exemplificative protocol of exposure to high levels of white light in albino rats is described. Then, the description of some main exemplificative outcomes deriving from the application of this protocol is provided, as well as practical suggestions used to assess the described topics.
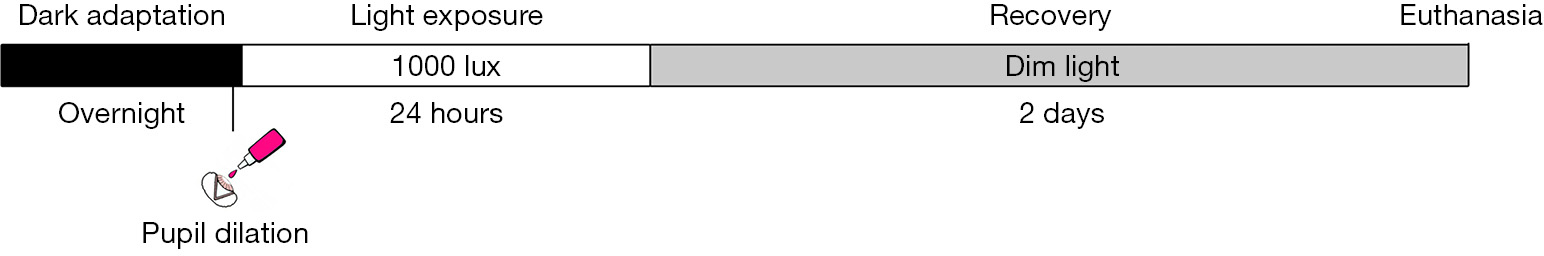
A schematic representation of this protocol is reported in Figure 2.
Retinal functional alterations following the experimental light paradigm can be evaluated by electroretinogram (ERG) from two days after light exposure. For instance, a full-field ERG can be performed to assess the activity of the entire retina in animals exposed to the intense light (12). The analysis could be performed non-invasively placing electrodes pairs on the corneal surface. Following a given light stimulus, positive and negative components of the recorded ERG wave indicate the summed activity of the potentials arising from different phases of retinal processing (12). ERG analyses should be performed under systemic anesthesia, although even a local anesthesia with eye drops, such as lidocaine, is advisable to prevent the animals from blinking. The dark-adapted scotopic ERG is one of the most widely used ERG analyses to evaluate retinal function (12). The recorded scotopic ERG wave comprises two main components. Indeed, after the light onset, an initial negative component (a-wave), corresponding to the photoreceptor contribution, is followed by a second positive-going b-wave from post-receptor retina (12). The considered light exposure protocol has been observed to reduce the amplitude of scotopic a- and b-waves of about 85–90% after LD (13-15). Moreover, longitudinal evaluation of LD-driven functional impairment during time revealed a partial recovery, maximal at 45 days, underlining a further deterioration after this time point, with no other recovery events (13,14).
The exposure to intense light induces biochemical changes in photoreceptors, resulting in cell damage and, consequently, death (16). The evaluation of photoreceptor death is one of the key features to analyze after a light exposure protocol, especially if a neuroprotective strategy is tested on these models. Such an evaluation could be performed at the molecular level, through the assessment of apoptotic markers’, upregulation, as well as by histological analyses which allow the quantification of dying cells among retinal layers. For example, the increase in pro-apoptotic molecules, as well as the activation of apoptotic-promoting caspases, could be measured by Western blot. The light exposure protocol has been reported to result in an increased pro-apoptotic Bax and cleaved caspase-3 levels even after 2 days of recovery in dim light (15). Then, the molecular analyses could be supported by immunohistochemical approaches, evaluating apoptotic-mediated DNA fragmentation by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay (14,15,17). In this respect, the increase in TUNEL positive cells could be detected even after 24 h of light exposure, then declining progressively within the first week of recovery in dim light, probably due to the progressive loss of cells (18,19). Photoreceptor degeneration occurring in this model is underlined ultimately by the thinning of outer nuclear layer (ONL) (16). Histological alterations in retinal layers after light exposure could be assessed by the labeling of retinal cell nuclei with 4',6-diamidino-2-phenylindole (DAPI) and the further quantification of ONL thickness on cross sections (15). Alternatively, the number of photoreceptor nuclei rows could be used as a measure of variations in ONL thickness (14). When evaluating the ONL thickness, the measure could be also expressed as the ratio between ONL to total retinal thickness, in order to account for putative oblique sectioning. In addition, the assessment of the thickness could be performed across either the entire retina or hemiretina (superior or inferior retina) by measuring across different eccentricities from the optic nerve (ON) and reporting the values as a function of the distance from ON (14). The light exposure protocol has been reported to induce the thinning of ONL, demonstrating a region-dependent loss of photoreceptors. Indeed, the most susceptible retinal region to LD is placed at the dorsal retina, in proximity of the so-called “hot-spot” (14,16). Unlike functional recovery, morphological alterations persisted during time (14).
One of the most important pathophysiological mechanisms determining photoreceptor death in LD is oxidative stress. Indeed, the excessive light exposure promotes a series of biochemical changes leading to the overproduction of ROS, which in turn can impair cell structure and function (4). In this context, ROS overproduction drives the activation of an endogenous antioxidant response, mediated by ROS-sensitive transcription factors, such as nuclear factor erythroid 2-related factor 2 (Nrf2), which in turn increase the production of antioxidant enzymes, such as heme oxygenase-1 (HO-1) (20,21). This occurs as a homeostatic response to counteract overproduction of ROS in the retina following light exposure. Therefore, to assess retinal oxidative stress it could be possible to either measure directly ROS levels through dedicated assays or, alternatively, evaluate putative changes in the levels of antioxidant markers as a response to increased ROS. In this respect, molecular analyses either at transcriptional or protein level could be useful to assess increments in antioxidant markers following LD. In the LD rats, protein levels of Nrf2, as well as HO-1, have been observed to increase even after 2 days of recovery in dim light after excessive light exposure (15).
LD protocol results in retinal gliosis and inflammation. The activation of retinal gliosis could be assessed through immunohistochemical analyses labeling retinal cross sections for reactive Müller cells [e.g., glial fibrillary acidic protein (GFAP)] and active microglial [e.g., ionized calcium-binding adapter molecule 1 (Iba1)] markers (15). Strong retinal upregulation of GFAP could be observed in LD model after 2 days of recovery in dim light, especially in Müller cells processes among retinal layers (15), further amplified during the first week of recovery (14,17). Moreover, the increase in GFAP fluorescence intensity was still observed even at later stages of recovery (14). Likewise, microglial activation, indicated by the increase in Iba1 positive cells, as well as the changed cell shape into active ameboid and the cell migration towards injured sites, was evidently detectable after 2 days of recovery and persisted until 60 days especially in dorsal retina (14,15). In addition, the activation of inflammatory processes, suggested by retinal gliosis and microglial activation, could be supported with molecular analysis of pro- and anti-inflammatory cytokines at transcriptional and protein levels. For example, qPCR, Western blot, and ELISA analyses suggested an upregulation of pro-inflammatory cytokines, such as interleukin 1 beta (IL-1β), interleukin 6 (IL-6) and tumor necrosis α (TNF-α), as well as a downregulation of anti-inflammatory molecules [e.g., interleukin 10 (IL-10)] after light exposure (15,18,19).
Retinal photoreceptor death is a feature common to different retinal diseases, such as retinitis pigmentosa and AMD (22). Therefore, the use of animal models mimicking photoreceptor degeneration could be fundamental to study the pathophysiological events and test the efficacy of neuroprotective treatments useful for both the diseases. The existence of many animal models of inherited retinal degeneration allows the study of cellular, biochemical, and molecular mechanisms responsible for photoreceptor death (7). However, the degeneration is rather slowly in these models and completes after several weeks to months. Thus, only a small number of cells are in the same dying stage throughout the degeneration, making the detection of subtle changes in levels, localization, and activity of involved molecules difficult (10).
It was demonstrated that an intense exposure to light can trigger oxidative stress, resulting in biochemical changes and ultimately leading to photoreceptor death (16). Therefore, it is possible to reproduce photoreceptor degeneration in animal models through the exposure of bright intense light. The pathophysiological processes observed in these models faithfully resemble the features of human retinal degenerations, making them a useful tool in studying disease mechanisms and treatments (8). The possibility to manipulate and control the severity of damage and the degeneration of photoreceptors, provides an advantage of LD lesions compared to genetical models. Indeed, the exposure to intense bright light can induce the synchronized activation of apoptosis in photoreceptors in a large area, which allows the evaluation of cellular and molecular events in a controlled fashion, facilitating the detection and the analysis (10). Moreover, protocols with various time exposures or light intensities can result in different severity of photoreceptor degeneration, possibly allowing a controlled gradeability of cell death (10). By varying the exposure parameters, such as the intensity of exposed light and the duration of the exposure, it is possible to obtain a certain degree of damage allowing the study of different features of the disease. For example, the exposure to different intensities of cold white light from 500 to 1,500 lux for 24 h has been observed to result in a dose-dependent loss of photoreceptors, with the superior retina displaying the most susceptibility to damage (23). Thus, the more intense the exposed light, the more severe the damage and the loss of photoreceptors might be. Furthermore, also modifying the time of exposure to a fixed light intensity might regulate the severity of damage. In this respect, an exposure to 1,000 lux of white light for 12 h has been observed to result in functional impairment and retinal gliosis, without any evident morphological changes and microglial activation. On the other hand, a longer exposure for 24 h at the same intensity has been demonstrated to induce even morphological alterations and microglial activation (14). Therefore, it is possible to adapt the exposure paradigm to the experimental needs, and the researcher has to evaluate what degree of damage should be obtained in order to study different mechanisms.
Despite their advantages, limitations of LD models which could influence the severity of damage should be considered. A series of variables must be carefully checked to establish the appropriate exposure paradigm able to reproduce the right intensity of damage. For instance, a higher susceptibility to LD was reported in older animals, demonstrating an age-dependent relationship (24). Moreover, also genetic background, strain differences and ocular pigmentation could influence the animal susceptibility to LD (10,24). Finally, an important attention should be paid on possible circadian variations in LD response. Indeed, LD susceptibility has been reported to depend on circadian variations (11). Thus, it is recommended to consider all of these variables in choosing the correct exposure paradigm, in order to have a protocol able to induce the needed effects without a high variability.
In conclusion, despite the variability introduced by these factors, LD models represent a useful tool to study the mechanisms responsible for photoreceptor degeneration. Thanks to the possibility to regulate the severity of damage and burst a synchronized apoptosis of photoreceptors, LD models are suitable to study degenerative mechanisms in a short time frame, in contrast to the longer times given by the slow-rated degenerative models. Nonetheless, it is still possible to adjust the exposure paradigm to obtain functional impairments without morphological outcomes, allowing the study of the early mechanisms behind LD-induced degeneration. Thus, it is important to choose the protocol according to the experimental needs, and the wide range of variables and limitations influencing the final outcomes should be taken into account to achieve proper results. In this respect, this study protocol seeks to provide basic guidelines of how a photo-oxidative damage is performed, using the described protocol as an example from the literature, and introduce the variables and limitations to give a critical overview of the factors to consider when choosing the right protocol. Thus, this study protocol should be considered as a starting tool for a critical beginning to the colleagues who want to approach with LD models, whose further details will be deepened by the literature of the chosen topic.









