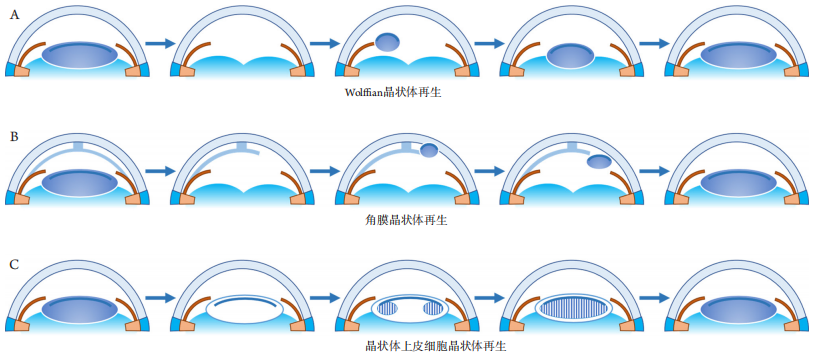
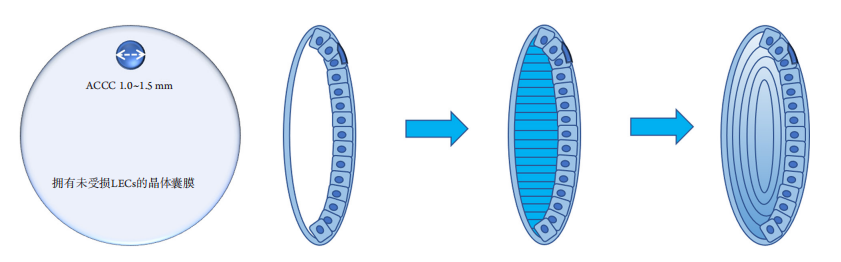
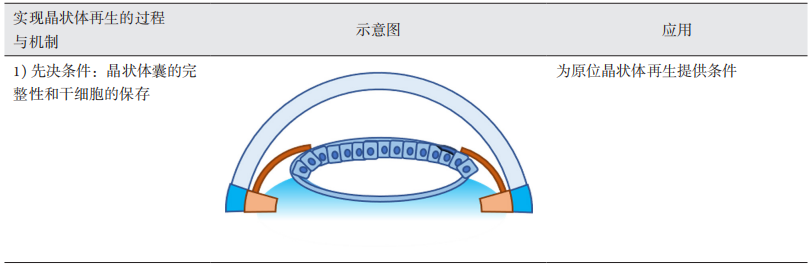
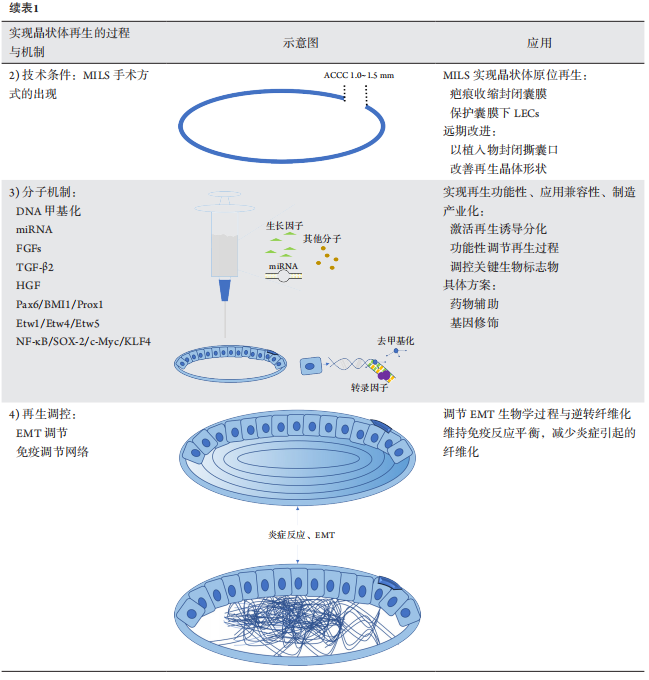
1、GBD 2019 Blindness and Vision Impairment Collaborators; Vision
Loss Expert Group of the Global Burden of Disease Study. Trends in
prevalence of blindness and distance and near vision impairment over
30 years: an analysis for the Global Burden of Disease Study[ J]. Lancet
Glob Health, 2021, 9(2): e130-e143.GBD 2019 Blindness and Vision Impairment Collaborators; Vision
Loss Expert Group of the Global Burden of Disease Study. Trends in
prevalence of blindness and distance and near vision impairment over
30 years: an analysis for the Global Burden of Disease Study[ J]. Lancet
Glob Health, 2021, 9(2): e130-e143.
2、GBD 2019 Blindness and Vision Impairment Collaborators; Vision
Loss Expert Group of the Global Burden of Disease Study. Causes of
blindness and vision impairment in 2020 and trends over 30 years,
and prevalence of avoidable blindness in relation to VISION 2020: the
Right to Sight: an analysis for the Global Burden of Disease Study[ J].
Lancet Glob Health, 2021, 9(2): e144-e160.GBD 2019 Blindness and Vision Impairment Collaborators; Vision
Loss Expert Group of the Global Burden of Disease Study. Causes of
blindness and vision impairment in 2020 and trends over 30 years,
and prevalence of avoidable blindness in relation to VISION 2020: the
Right to Sight: an analysis for the Global Burden of Disease Study[ J].
Lancet Glob Health, 2021, 9(2): e144-e160.
3、Xia H, Li X, Gao W, et al. Tissue repair and regeneration with
endogenous stem cells[ J]. Nat Rev Mater, 2018, 3(7): 174-193.Xia H, Li X, Gao W, et al. Tissue repair and regeneration with
endogenous stem cells[ J]. Nat Rev Mater, 2018, 3(7): 174-193.
4、Stern JH, Tian Y, Funderburgh J, et al. Regenerating eye tissues to
preserve and restore vision[ J]. Cell Stem Cell, 2018, 22(6): 834-849.Stern JH, Tian Y, Funderburgh J, et al. Regenerating eye tissues to
preserve and restore vision[ J]. Cell Stem Cell, 2018, 22(6): 834-849.
5、Dietz AB, Dozois EJ, Fletcher JG, et al. Autologous mesenchymal stem
cells, applied in a bioabsorbable matrix, for treatment of perianal fistulas
in patients with Crohn's disease[ J]. Gastroenterology, 2017, 153(1):
59-62.e2.Dietz AB, Dozois EJ, Fletcher JG, et al. Autologous mesenchymal stem
cells, applied in a bioabsorbable matrix, for treatment of perianal fistulas
in patients with Crohn's disease[ J]. Gastroenterology, 2017, 153(1):
59-62.e2.
6、Eichstadt S, Barriga M, Ponakala A, et al. Phase 1/2a clinical trial
of gene-corrected autologous cell therapy for recessive dystrophic
epidermolysis bullosa[ J]. JCI Insight, 2019, 4(19): 130554.Eichstadt S, Barriga M, Ponakala A, et al. Phase 1/2a clinical trial
of gene-corrected autologous cell therapy for recessive dystrophic
epidermolysis bullosa[ J]. JCI Insight, 2019, 4(19): 130554.
7、Gonzalez G, Sasamoto Y, Ksander BR, et al. Limbal stem cells: identity,
developmental origin, and therapeutic potential[ J]. Wiley Interdiscip
Rev Dev Biol, 2018, 7(2): 10.1002/wdev.303.Gonzalez G, Sasamoto Y, Ksander BR, et al. Limbal stem cells: identity,
developmental origin, and therapeutic potential[ J]. Wiley Interdiscip
Rev Dev Biol, 2018, 7(2): 10.1002/wdev.303.
8、Jiang S, Tian G, Li X, et al. Research progress on stem cell therapies
for articular cartilage regeneration[ J]. Stem Cells Int, 2021, 2021:
8882505.Jiang S, Tian G, Li X, et al. Research progress on stem cell therapies
for articular cartilage regeneration[ J]. Stem Cells Int, 2021, 2021:
8882505.
9、Henry JJ, Hamilton PW. Diverse evolutionary origins and mechanisms
of lens regeneration[ J]. Mol Biol Evol, 2018, 35(7): 1563-1575.Henry JJ, Hamilton PW. Diverse evolutionary origins and mechanisms
of lens regeneration[ J]. Mol Biol Evol, 2018, 35(7): 1563-1575.
10、Sukhija J, Kaur S. Nature nurtures: lens regeneration, a breakthrough in
ophthalmology[ J]. Ann Eye Sci, 2017, 2: 17.Sukhija J, Kaur S. Nature nurtures: lens regeneration, a breakthrough in
ophthalmology[ J]. Ann Eye Sci, 2017, 2: 17.
11、Wan J, Goldman D. Retina regeneration in zebrafish[ J]. Curr Opin
Genet Dev, 2016, 40: 41-47.Wan J, Goldman D. Retina regeneration in zebrafish[ J]. Curr Opin
Genet Dev, 2016, 40: 41-47.
12、Brockes JP. Amphibian limb regeneration: rebuilding a complex
structure[ J]. Science, 1997, 276(5309): 81-87.Brockes JP. Amphibian limb regeneration: rebuilding a complex
structure[ J]. Science, 1997, 276(5309): 81-87.
13、Goldman D. Müller glial cell reprogramming and retina regeneration[ J].
Nat Rev Neurosci, 2014, 15(7): 431-442.Goldman D. Müller glial cell reprogramming and retina regeneration[ J].
Nat Rev Neurosci, 2014, 15(7): 431-442.
14、Henry JJ, Tsonis PA. Molecular and cellular aspects of amphibian lens
regeneration[ J]. Prog Retin Eye Res, 2010, 29(6): 543-555.Henry JJ, Tsonis PA. Molecular and cellular aspects of amphibian lens
regeneration[ J]. Prog Retin Eye Res, 2010, 29(6): 543-555.
15、Wolff G. Entwickelungsphysiologische studien[ J]. Arch Entwicklmech
Org, 1895, 1(3): 380-390.Wolff G. Entwickelungsphysiologische studien[ J]. Arch Entwicklmech
Org, 1895, 1(3): 380-390.
16、Ledwon JK, Turin SY, Gosain AK, et al. The expression of fgfr3 in the
zebrafish head[ J]. Gene Expr Patterns, 2018, 29: 32-38.Ledwon JK, Turin SY, Gosain AK, et al. The expression of fgfr3 in the
zebrafish head[ J]. Gene Expr Patterns, 2018, 29: 32-38.
17、Filoni S, Bosco L, Cioni C. Reconstruction of the crystalline lens from
fragments of capsular membrane and epithelium in larvae of Rana
esculenta[ J]. Acta Embryol Exp (Palermo), 1977, (1): 41-49.Filoni S, Bosco L, Cioni C. Reconstruction of the crystalline lens from
fragments of capsular membrane and epithelium in larvae of Rana
esculenta[ J]. Acta Embryol Exp (Palermo), 1977, (1): 41-49.
18、Tsonis PA. Lens regeneration[M]//Encyclopedia of the eye. Oxford:
Academic Press, 2010: 557-564.Tsonis PA. Lens regeneration[M]//Encyclopedia of the eye. Oxford:
Academic Press, 2010: 557-564.
19、Perry KJ, Thomas AG, Henry JJ. Expression of pluripotency factors in
larval epithelia of the frog Xenopus: evidence for the presence of cornea
epithelial stem cells[ J]. Dev Biol, 2013, 374(2): 281-294.Perry KJ, Thomas AG, Henry JJ. Expression of pluripotency factors in
larval epithelia of the frog Xenopus: evidence for the presence of cornea
epithelial stem cells[ J]. Dev Biol, 2013, 374(2): 281-294.
20、Perry KJ, Hamilton PW, Sonam S, et al. The role of sensory innervation
in cornea-lens regeneration[ J]. Dev Dyn, 2019, 248(7): 530-544.Perry KJ, Hamilton PW, Sonam S, et al. The role of sensory innervation
in cornea-lens regeneration[ J]. Dev Dyn, 2019, 248(7): 530-544.
21、刘晓敏, 代云海, 谢立信. 哺乳动物晶状体再生的研究进展[ J].
中华眼科杂志, 2019, 55(7): 549-553.
LIU Xiaomin, DAI Yunhai, XIE Lixin. Advances and clinical application
of lens regeneration in mammal[ J]. Chinese Journal of Ophthalmology,
2019, 55(7): 549-553.刘晓敏, 代云海, 谢立信. 哺乳动物晶状体再生的研究进展[ J].
中华眼科杂志, 2019, 55(7): 549-553.
LIU Xiaomin, DAI Yunhai, XIE Lixin. Advances and clinical application
of lens regeneration in mammal[ J]. Chinese Journal of Ophthalmology,
2019, 55(7): 549-553.
22、Zhou KJ, Li YN, Huang FR , et al. In vivo observation of lens
regeneration in rat using ultra-long scan depth optical coherence
tomography[ J]. Invest Ophthalmol Vis Sci, 2016, 57(15): 6615-6623.Zhou KJ, Li YN, Huang FR , et al. In vivo observation of lens
regeneration in rat using ultra-long scan depth optical coherence
tomography[ J]. Invest Ophthalmol Vis Sci, 2016, 57(15): 6615-6623.
23、Zukin LM, Pedler MG, Chyung K, et al. Aldose reductase inhibition
enhances lens regeneration in mice[ J]. Chem Biol Interact, 2019, 307:
58-62.Zukin LM, Pedler MG, Chyung K, et al. Aldose reductase inhibition
enhances lens regeneration in mice[ J]. Chem Biol Interact, 2019, 307:
58-62.
24、柳夏林,张新愉,刘奕志,等. 兔眼晶状体再生模型的建立及观
察[ J]. 眼科学报, 2002, 18(4): 230-234.
LIU Xialin, ZHANG Xinyu, LIU Yizhi, et al. To establish and observe
the experimental lens regeneration model in rabbits[ J]. Yan Ke Xue
Bao, 2002, 18(4): 230-234.柳夏林,张新愉,刘奕志,等. 兔眼晶状体再生模型的建立及观
察[ J]. 眼科学报, 2002, 18(4): 230-234.
LIU Xialin, ZHANG Xinyu, LIU Yizhi, et al. To establish and observe
the experimental lens regeneration model in rabbits[ J]. Yan Ke Xue
Bao, 2002, 18(4): 230-234.
25、Andjeli? S, Dra?lar K, Lumi X, et al. Morphological and proliferative
studies on ex vivo cultured human anterior lens epithelial cells -
relevance to capsular opacification[ J]. Acta Ophthalmol, 2015, 93(6):
e499-e506.Andjeli? S, Dra?lar K, Lumi X, et al. Morphological and proliferative
studies on ex vivo cultured human anterior lens epithelial cells -
relevance to capsular opacification[ J]. Acta Ophthalmol, 2015, 93(6):
e499-e506.
26、Lin H, Ouyang H, Zhu J, et al. Lens regeneration using endogenous
stem cells with gain of visual function[ J]. Nature, 2016, 531(7594):
323-328.Lin H, Ouyang H, Zhu J, et al. Lens regeneration using endogenous
stem cells with gain of visual function[ J]. Nature, 2016, 531(7594):
323-328.
27、Han C, Li J, Wang C, et al. Wnt5a contributes to the differentiation
of human embryonic stem cells into lentoid bodies through the
noncanonical Wnt/JNK signaling pathway[ J]. Invest Ophthalmol Vis
Sci, 2018, 59(8): 3449-3460.Han C, Li J, Wang C, et al. Wnt5a contributes to the differentiation
of human embryonic stem cells into lentoid bodies through the
noncanonical Wnt/JNK signaling pathway[ J]. Invest Ophthalmol Vis
Sci, 2018, 59(8): 3449-3460.
28、Anchan RM, Lachke SA, Gerami-Naini B, et al. Pax6- and Six3-
mediated induction of lens cell fate in mouse and human ES cells[ J].
PLoS One, 2014, 9(12): e115106.Anchan RM, Lachke SA, Gerami-Naini B, et al. Pax6- and Six3-
mediated induction of lens cell fate in mouse and human ES cells[ J].
PLoS One, 2014, 9(12): e115106.
29、Gwon A . Lens regeneration in mammals: a rev iew[ J]. Sur v
Ophthalmol, 2006, 51(1): 51-62.Gwon A . Lens regeneration in mammals: a rev iew[ J]. Sur v
Ophthalmol, 2006, 51(1): 51-62.
30、Nagamoto T, Tanaka N, Fujiwara T. Inhibition of posterior capsule
opacification by a capsular adhesion-preventing ring[ J]. Arch
Ophthalmol, 2009, 127(4): 471-474.Nagamoto T, Tanaka N, Fujiwara T. Inhibition of posterior capsule
opacification by a capsular adhesion-preventing ring[ J]. Arch
Ophthalmol, 2009, 127(4): 471-474.
31、Shammas HJ. Relaxing the fibrosed capsulorhexis rim to correct
induced hyperopia after phacoemulsification[ J]. J Cataract Refract
Surg, 1995, 21(2): 228-229.Shammas HJ. Relaxing the fibrosed capsulorhexis rim to correct
induced hyperopia after phacoemulsification[ J]. J Cataract Refract
Surg, 1995, 21(2): 228-229.
32、Martínez Toldos JJ, Artola Roig A, Chipont Benabent E. Total anterior
capsule closure after silicone intraocular lens implantation[ J]. J
Cataract Refract Surg, 1996, 22(2): 269-271.Martínez Toldos JJ, Artola Roig A, Chipont Benabent E. Total anterior
capsule closure after silicone intraocular lens implantation[ J]. J
Cataract Refract Surg, 1996, 22(2): 269-271.
33、Hurvitz LM. YAG anterior capsulectomy and lysis of posterior
synechiae after cataract surgery[ J]. Ophthalmic Surg, 1992, 23(2):
103-107.Hurvitz LM. YAG anterior capsulectomy and lysis of posterior
synechiae after cataract surgery[ J]. Ophthalmic Surg, 1992, 23(2):
103-107.
34、Gwon A , Gruber L. Engineering the cr ystalline lens w ith a
biodegradable or non-degradable scaffold[ J]. Exp Eye Res, 2010,
91(2): 220-228.Gwon A , Gruber L. Engineering the cr ystalline lens w ith a
biodegradable or non-degradable scaffold[ J]. Exp Eye Res, 2010,
91(2): 220-228.
35、Tan X, Zhu Y, Chen C, et al. Sprouty2 suppresses epithelial-
mesenchymal transition of human lens epithelial cells through
blockade of Smad2 and ERK1/2 pathways[ J]. PLoS One, 2016, 11(7):
e0159275.Tan X, Zhu Y, Chen C, et al. Sprouty2 suppresses epithelial-
mesenchymal transition of human lens epithelial cells through
blockade of Smad2 and ERK1/2 pathways[ J]. PLoS One, 2016, 11(7):
e0159275.
36、Jiang J, Shihan MH, Wang Y, et al. Lens epithelial cells initiate
an inflammatory response following cataract surgery[ J]. Invest
Ophthalmol Vis Sci, 2018, 59(12): 4986-4997.Jiang J, Shihan MH, Wang Y, et al. Lens epithelial cells initiate
an inflammatory response following cataract surgery[ J]. Invest
Ophthalmol Vis Sci, 2018, 59(12): 4986-4997.
37、Inanc M, Tekin K, Erol YO, et al. The ultrastructural alterations in the
lens capsule and epithelium in eyes with traumatic white cataract[ J].
Int Ophthalmol, 2019, 39(1): 47-53.Inanc M, Tekin K, Erol YO, et al. The ultrastructural alterations in the
lens capsule and epithelium in eyes with traumatic white cataract[ J].
Int Ophthalmol, 2019, 39(1): 47-53.
38、Han Y, Tang J, Xia J, et al. Anti-adhesive and antiproliferative synergistic
surface modification of intraocular lens for reduced posterior capsular
opacification[ J]. Int J Nanomedicine, 2019, 14: 9047-9061.Han Y, Tang J, Xia J, et al. Anti-adhesive and antiproliferative synergistic
surface modification of intraocular lens for reduced posterior capsular
opacification[ J]. Int J Nanomedicine, 2019, 14: 9047-9061.
39、Lin L, Lin Q, Li J, et al. ROCK inhibitor modified intraocular lens as an
approach for inhibiting the proliferation and migration of lens epithelial
cells and posterior capsule opacification[ J]. Biomater Sci, 2019, 7(10):
4208-4217.Lin L, Lin Q, Li J, et al. ROCK inhibitor modified intraocular lens as an
approach for inhibiting the proliferation and migration of lens epithelial
cells and posterior capsule opacification[ J]. Biomater Sci, 2019, 7(10):
4208-4217.
40、Huang Y, Xie L. Expression of transcription factors and crystallin
proteins during rat lens regeneration[ J]. Mol Vis, 2010, 16: 341-352.Huang Y, Xie L. Expression of transcription factors and crystallin
proteins during rat lens regeneration[ J]. Mol Vis, 2010, 16: 341-352.
41、Wu X, Liu Z, Zhang X, et al. Proteomics analysis and proteogenomic
characterization of different physiopathological human lenses[ J]. BMC
Ophthalmol, 2017, 17(1): 253.Wu X, Liu Z, Zhang X, et al. Proteomics analysis and proteogenomic
characterization of different physiopathological human lenses[ J]. BMC
Ophthalmol, 2017, 17(1): 253.
42、Khurana I, Al-Hasani K, Maxwell S, et al. DNA methylation status
correlates with adult β-cell regeneration capacity[ J]. NPJ Regen Med,
2021, 6(1): 7.Khurana I, Al-Hasani K, Maxwell S, et al. DNA methylation status
correlates with adult β-cell regeneration capacity[ J]. NPJ Regen Med,
2021, 6(1): 7.
43、Peng S, Shi S, Tao G, et al. JKAMP inhibits the osteogenic capacity of
adipose-derived stem cells in diabetic osteoporosis by modulating the
Wnt signaling pathway through intragenic DNA methylation[ J]. Stem
Cell Res Ther, 2021, 12(1): 120.Peng S, Shi S, Tao G, et al. JKAMP inhibits the osteogenic capacity of
adipose-derived stem cells in diabetic osteoporosis by modulating the
Wnt signaling pathway through intragenic DNA methylation[ J]. Stem
Cell Res Ther, 2021, 12(1): 120.
44、Li R, Li B, Cao Y, et al. Long non-coding RNA Mir22hg-derived miR-
22-3p promotes skeletal muscle differentiation and regeneration by
inhibiting HDAC4[ J]. Mol Ther Nucleic Acids, 2021, 24: 200-211.Li R, Li B, Cao Y, et al. Long non-coding RNA Mir22hg-derived miR-
22-3p promotes skeletal muscle differentiation and regeneration by
inhibiting HDAC4[ J]. Mol Ther Nucleic Acids, 2021, 24: 200-211.
45、Sevilla A, Papatsenko D, Mazloom AR, et al. An Esrrb and Nanog cell
fate regulatory module controlled by feed forward loop interactions[ J].
Front Cell Dev Biol, 2021, 9: 630067.Sevilla A, Papatsenko D, Mazloom AR, et al. An Esrrb and Nanog cell
fate regulatory module controlled by feed forward loop interactions[ J].
Front Cell Dev Biol, 2021, 9: 630067.
46、Wang Y, Guan H. The Role of DNA methylation in lens development
and cataract formation[ J]. Cell Mol Neurobiol, 2017, 37(6): 979-984.Wang Y, Guan H. The Role of DNA methylation in lens development
and cataract formation[ J]. Cell Mol Neurobiol, 2017, 37(6): 979-984.
47、Lu Y, Brommer B, Tian X, et al. Reprogramming to recover youthful
epigenetic information and restore vision[ J]. Nature, 2020, 588(7836):
124-129.Lu Y, Brommer B, Tian X, et al. Reprogramming to recover youthful
epigenetic information and restore vision[ J]. Nature, 2020, 588(7836):
124-129.
48、Dirks RP, Klok EJ, van Genesen ST, et al. The sequence of regulatory
events controlling the expression of the gamma D-crystallin gene during
fibroblast growth factor-mediated rat lens fiber cell differentiation[ J].
Dev Biol, 1996, 173(1): 14-25.Dirks RP, Klok EJ, van Genesen ST, et al. The sequence of regulatory
events controlling the expression of the gamma D-crystallin gene during
fibroblast growth factor-mediated rat lens fiber cell differentiation[ J].
Dev Biol, 1996, 173(1): 14-25.
49、Chen X, Xiao W, Chen W, et al. MicroRNA-26a and -26b inhibit lens
fibrosis and cataract by negatively regulating Jagged-1/Notch signaling
pathway[ J]. Cell Death Differ, 2017, 24(8): 1431-1442.Chen X, Xiao W, Chen W, et al. MicroRNA-26a and -26b inhibit lens
fibrosis and cataract by negatively regulating Jagged-1/Notch signaling
pathway[ J]. Cell Death Differ, 2017, 24(8): 1431-1442.
50、McAvoy JW, Chamberlain CG. Fibroblast growth factor (FGF)
induces different responses in lens epithelial cells depending on its
concentration[ J]. Development, 1989, 107(2): 221-228.McAvoy JW, Chamberlain CG. Fibroblast growth factor (FGF)
induces different responses in lens epithelial cells depending on its
concentration[ J]. Development, 1989, 107(2): 221-228.
51、Shu DY, Lovicu FJ. Enhanced EGF receptor-signaling potentiates
TGFβ-induced lens epithelial-mesenchymal transition[ J]. Exp Eye Res,
2019, 185: 107693.Shu DY, Lovicu FJ. Enhanced EGF receptor-signaling potentiates
TGFβ-induced lens epithelial-mesenchymal transition[ J]. Exp Eye Res,
2019, 185: 107693.
52、Huang X, Wang Y, Zhang P, et al. A HGF-derived peptide suppresses
EMT in human lens epithelial cells via the TGF-β/Smad and Akt/
mTOR signaling pathways[ J]. Mol Med Rep, 2020, 22(1): 551-558.Huang X, Wang Y, Zhang P, et al. A HGF-derived peptide suppresses
EMT in human lens epithelial cells via the TGF-β/Smad and Akt/
mTOR signaling pathways[ J]. Mol Med Rep, 2020, 22(1): 551-558.
53、Aryal S, Viet J, Weatherbee BAT, et al. The cataract-linked RNA-binding
protein Celf1 post-transcriptionally controls the spatiotemporal
expression of the key homeodomain transcription factors Pax6 and
Prox1 in lens development[ J]. Hum Genet, 2020, 139(12): 1541-1554.Aryal S, Viet J, Weatherbee BAT, et al. The cataract-linked RNA-binding
protein Celf1 post-transcriptionally controls the spatiotemporal
expression of the key homeodomain transcription factors Pax6 and
Prox1 in lens development[ J]. Hum Genet, 2020, 139(12): 1541-1554.
54、Garg A, Hannan A, Wang Q, et al. Etv transcription factors functionally
diverge from their upstream FGF signaling in lens development[ J].
Elife, 2020, 9: 51915.Garg A, Hannan A, Wang Q, et al. Etv transcription factors functionally
diverge from their upstream FGF signaling in lens development[ J].
Elife, 2020, 9: 51915.
55、Sun Y, Rong X, Li D, et al. NF-κB/cartilage acidic protein 1 promotes
ultraviolet B irradiation-induced apoptosis of human lens epithelial
cells[ J]. DNA Cell Biol, 2020, 39(4): 513-521.Sun Y, Rong X, Li D, et al. NF-κB/cartilage acidic protein 1 promotes
ultraviolet B irradiation-induced apoptosis of human lens epithelial
cells[ J]. DNA Cell Biol, 2020, 39(4): 513-521.
56、Maki N, Suetsugu-Maki R, Tarui H, et al. Expression of stem cell
pluripotency factors during regeneration in newts[ J]. Dev Dyn, 2009,
238(6): 1613-1616.Maki N, Suetsugu-Maki R, Tarui H, et al. Expression of stem cell
pluripotency factors during regeneration in newts[ J]. Dev Dyn, 2009,
238(6): 1613-1616.
57、Zhao Y, Zheng D, Cvekl A. Profiling of chromatin accessibility and
identification of general cis-regulatory mechanisms that control two
ocular lens differentiation pathways[ J]. Epigenetics Chromatin, 2019,
12(1): 27.Zhao Y, Zheng D, Cvekl A. Profiling of chromatin accessibility and
identification of general cis-regulatory mechanisms that control two
ocular lens differentiation pathways[ J]. Epigenetics Chromatin, 2019,
12(1): 27.
58、Shaw TJ, Martin P. Wound repair: a showcase for cell plasticity and
migration[ J]. Curr Opin Cell Biol, 2016, 42: 29-37.Shaw TJ, Martin P. Wound repair: a showcase for cell plasticity and
migration[ J]. Curr Opin Cell Biol, 2016, 42: 29-37.
59、Arnoux V, Nassour M, L'Helgoualc'h A, et al. Erk5 controls Slug
expression and keratinocyte activation during wound healing[ J]. Mol
Biol Cell, 2008, 19(11): 4738-4749.Arnoux V, Nassour M, L'Helgoualc'h A, et al. Erk5 controls Slug
expression and keratinocyte activation during wound healing[ J]. Mol
Biol Cell, 2008, 19(11): 4738-4749.
60、Aurora AB, Olson EN. Immune modulation of stem cells and
regeneration[ J]. Cell Stem Cell, 2014, 15(1): 14-25.Aurora AB, Olson EN. Immune modulation of stem cells and
regeneration[ J]. Cell Stem Cell, 2014, 15(1): 14-25.
61、Zhou R, Caspi RR. Ocular immune privilege[ J]. F1000 Biol Rep,
2010, 2: 3.Zhou R, Caspi RR. Ocular immune privilege[ J]. F1000 Biol Rep,
2010, 2: 3.
62、Spierer A, Desatnik H, Blumenthal M. Refractive status in children
after long-term follow up of cataract surgery with intraocular lens
implantation[ J]. J Pediatr Ophthalmol Strabismus, 1999, 36(1): 25-29.Spierer A, Desatnik H, Blumenthal M. Refractive status in children
after long-term follow up of cataract surgery with intraocular lens
implantation[ J]. J Pediatr Ophthalmol Strabismus, 1999, 36(1): 25-29.
63、Ursell PG, Dhariwal M, O'Boyle D, et al. 5-year incidence of YAG
capsulotomy and PCO after cataract surgery with single-piece
monofocal intraocular lenses: a real-world evidence study of 20,763
eyes[ J]. Eye (Lond), 2020, 34(5): 960-968.Ursell PG, Dhariwal M, O'Boyle D, et al. 5-year incidence of YAG
capsulotomy and PCO after cataract surgery with single-piece
monofocal intraocular lenses: a real-world evidence study of 20,763
eyes[ J]. Eye (Lond), 2020, 34(5): 960-968.
64、Logan CM, Bowen CJ, Menko AS. Induction of immune surveillance of
the dysmorphogenic lens[ J]. Sci Rep, 2017, 7(1): 16235.Logan CM, Bowen CJ, Menko AS. Induction of immune surveillance of
the dysmorphogenic lens[ J]. Sci Rep, 2017, 7(1): 16235.
65、Malloch EL, Perry KJ, Fukui L, et al. Gene expression profiles of lens
regeneration and development in Xenopus laevis[ J]. Dev Dyn, 2009,
238(9): 2340-2356.Malloch EL, Perry KJ, Fukui L, et al. Gene expression profiles of lens
regeneration and development in Xenopus laevis[ J]. Dev Dyn, 2009,
238(9): 2340-2356.
66、Brockes JP, Kumar A. Comparative aspects of animal regeneration[ J].
Annu Rev Cell Dev Biol, 2008, 24: 525-549.Brockes JP, Kumar A. Comparative aspects of animal regeneration[ J].
Annu Rev Cell Dev Biol, 2008, 24: 525-549.
67、Qin C, Liu S, Wen S, et al. Enhanced PCO prevention of drug eluting
IOLs via endocytosis and autophagy effects of a PAMAM dendrimer[ J].
J Mater Chem B, 2021, 9(3): 793-800.Qin C, Liu S, Wen S, et al. Enhanced PCO prevention of drug eluting
IOLs via endocytosis and autophagy effects of a PAMAM dendrimer[ J].
J Mater Chem B, 2021, 9(3): 793-800.
68、Huang H, Zhu S, Liu D, et al. Antiproliferative drug-loaded multi-
functionalized intraocular lens for reducing posterior capsular
opacification[ J]. J Biomater Sci Polym Ed, 2021, 32(6): 735-748.
Huang H, Zhu S, Liu D, et al. Antiproliferative drug-loaded multi-
functionalized intraocular lens for reducing posterior capsular
opacification[ J]. J Biomater Sci Polym Ed, 2021, 32(6): 735-748.
69、Tang J, Liu S, Han Y, et al. Surface modification of intraocular lenses
via photodynamic coating for safe and effective PCO prevention[ J]. J
Mater Chem B, 2021, 9(6): 1546-1556.Tang J, Liu S, Han Y, et al. Surface modification of intraocular lenses
via photodynamic coating for safe and effective PCO prevention[ J]. J
Mater Chem B, 2021, 9(6): 1546-1556.
70、Vannella KM, Wynn TA. Mechanisms of organ injury and repair by
macrophages[ J]. Annu Rev Physiol, 2017, 79: 593-617.Vannella KM, Wynn TA. Mechanisms of organ injury and repair by
macrophages[ J]. Annu Rev Physiol, 2017, 79: 593-617.
71、Marote A, Teixeira FG, Mendes-Pinheiro B, et al. MSCs-derived
exosomes: cell-secreted nanovesicles with regenerative potential[ J].
Front Pharmacol, 2016, 7: 231.Marote A, Teixeira FG, Mendes-Pinheiro B, et al. MSCs-derived
exosomes: cell-secreted nanovesicles with regenerative potential[ J].
Front Pharmacol, 2016, 7: 231.
72、Ueki Y, Wilken MS, Cox KE, et al. Transgenic expression of the
proneural transcription factor Ascl1 in Müller glia stimulates retinal
regeneration in young mice[ J]. Proc Natl Acad Sci U S A, 2015,
112(44): 13717-13722.Ueki Y, Wilken MS, Cox KE, et al. Transgenic expression of the
proneural transcription factor Ascl1 in Müller glia stimulates retinal
regeneration in young mice[ J]. Proc Natl Acad Sci U S A, 2015,
112(44): 13717-13722.
73、Yao K, Qiu S, Wang YV, et al. Restoration of vision after de novo
genesis of rod photoreceptors in mammalian retinas[ J]. Nature, 2018,
560(7719): 484-488.Yao K, Qiu S, Wang YV, et al. Restoration of vision after de novo
genesis of rod photoreceptors in mammalian retinas[ J]. Nature, 2018,
560(7719): 484-488.
74、Amano S, Yamagami S, Mimura T, et al. Corneal stromal and
endothelial cell precursors[ J]. Cornea, 2006, 25(10 Suppl 1): S73-S77.Amano S, Yamagami S, Mimura T, et al. Corneal stromal and
endothelial cell precursors[ J]. Cornea, 2006, 25(10 Suppl 1): S73-S77.
75、Koizumi N, Okumura N, Ueno M, et al. New therapeutic modality for
corneal endothelial disease using Rho-associated kinase inhibitor eye
drops[ J]. Cornea, 2014, 33 Suppl 11: S25-S31.Koizumi N, Okumura N, Ueno M, et al. New therapeutic modality for
corneal endothelial disease using Rho-associated kinase inhibitor eye
drops[ J]. Cornea, 2014, 33 Suppl 11: S25-S31.
76、Wang EX, Jiang X. Stem cells from trabecular meshwork cells can
secrete extracellular matrix[ J]. Biochem Biophys Res Commun, 2020,
523(2): 522-526.Wang EX, Jiang X. Stem cells from trabecular meshwork cells can
secrete extracellular matrix[ J]. Biochem Biophys Res Commun, 2020,
523(2): 522-526.
77、Donati G, Watt FM. Stem cell heterogeneity and plasticity in
epithelia[ J]. Cell Stem Cell, 2015, 16(5): 465-476.Donati G, Watt FM. Stem cell heterogeneity and plasticity in
epithelia[ J]. Cell Stem Cell, 2015, 16(5): 465-476.
78、van der Flier LG, Clevers H. Stem cells, self-renewal, and differentiation
in the intestinal epithelium[ J]. Annu Rev Physiol, 2009, 71: 241-260.van der Flier LG, Clevers H. Stem cells, self-renewal, and differentiation
in the intestinal epithelium[ J]. Annu Rev Physiol, 2009, 71: 241-260.
79、Servick K. Stem cell approach for cataracts challenged[ J]. Science,
2017, 356(6345): 1318-1319.Servick K. Stem cell approach for cataracts challenged[ J]. Science,
2017, 356(6345): 1318-1319.
80、Laha B, Stafford BK, Huberman AD. Regenerating optic pathways from
the eye to the brain[ J]. Science, 2017, 356(6342): 1031-1034.Laha B, Stafford BK, Huberman AD. Regenerating optic pathways from
the eye to the brain[ J]. Science, 2017, 356(6342): 1031-1034.
81、Kinoshita S, Koizumi N, Ueno M, et al. Injection of cultured cells
with a rock inhibitor for bullous keratopathy[ J]. N Engl J Med, 2018,
378(11): 995-1003.Kinoshita S, Koizumi N, Ueno M, et al. Injection of cultured cells
with a rock inhibitor for bullous keratopathy[ J]. N Engl J Med, 2018,
378(11): 995-1003.
82、Karin M, Clevers H. Reparative inflammation takes charge of tissue
regeneration[ J]. Nature, 2016, 529(7586): 307-315.Karin M, Clevers H. Reparative inflammation takes charge of tissue
regeneration[ J]. Nature, 2016, 529(7586): 307-315.
83、Leeman DS, Hebestreit K, Ruetz T, et al. Lysosome activation clears
aggregates and enhances quiescent neural stem cell activation during
aging[ J]. Science, 2018, 359(6381): 1277-1283.Leeman DS, Hebestreit K, Ruetz T, et al. Lysosome activation clears
aggregates and enhances quiescent neural stem cell activation during
aging[ J]. Science, 2018, 359(6381): 1277-1283.
84、Perico L, Morigi M, Rota C, et al. Human mesenchymal stromal
cells transplanted into mice stimulate renal tubular cells and enhance
mitochondrial function[ J]. Nat Commun, 2017, 8(1): 983.Perico L, Morigi M, Rota C, et al. Human mesenchymal stromal
cells transplanted into mice stimulate renal tubular cells and enhance
mitochondrial function[ J]. Nat Commun, 2017, 8(1): 983.
85、Otsuki L, Brand AH. Cell cycle heterogeneity directs the timing
of neural stem cell activation from quiescence[ J]. Science, 2018,
360(6384): 99-102.Otsuki L, Brand AH. Cell cycle heterogeneity directs the timing
of neural stem cell activation from quiescence[ J]. Science, 2018,
360(6384): 99-102.
86、Bassat E, Mutlak YE, Genzelinakh A, et al. The extracellular matrix
protein agrin promotes heart regeneration in mice[ J]. Nature, 2017,
547(7662): 179-184.Bassat E, Mutlak YE, Genzelinakh A, et al. The extracellular matrix
protein agrin promotes heart regeneration in mice[ J]. Nature, 2017,
547(7662): 179-184.
87、Zhang Y, Kim MS, Jia B, et al. Hypothalamic stem cells control ageing
speed partly through exosomal miRNAs[ J]. Nature, 2017, 548(7665):
52-57.Zhang Y, Kim MS, Jia B, et al. Hypothalamic stem cells control ageing
speed partly through exosomal miRNAs[ J]. Nature, 2017, 548(7665):
52-57.
88、Jorstad NL, Wilken MS, Grimes WN, et al. Stimulation of functional
neuronal regeneration from Müller glia in adult mice[ J]. Nature, 2017,
548(7665): 103-107.Jorstad NL, Wilken MS, Grimes WN, et al. Stimulation of functional
neuronal regeneration from Müller glia in adult mice[ J]. Nature, 2017,
548(7665): 103-107.
89、Tzahor E, Poss KD. Cardiac regeneration strategies: staying young at
heart[ J]. Science, 2017, 356(6342): 1035-1039.Tzahor E, Poss KD. Cardiac regeneration strategies: staying young at
heart[ J]. Science, 2017, 356(6342): 1035-1039.