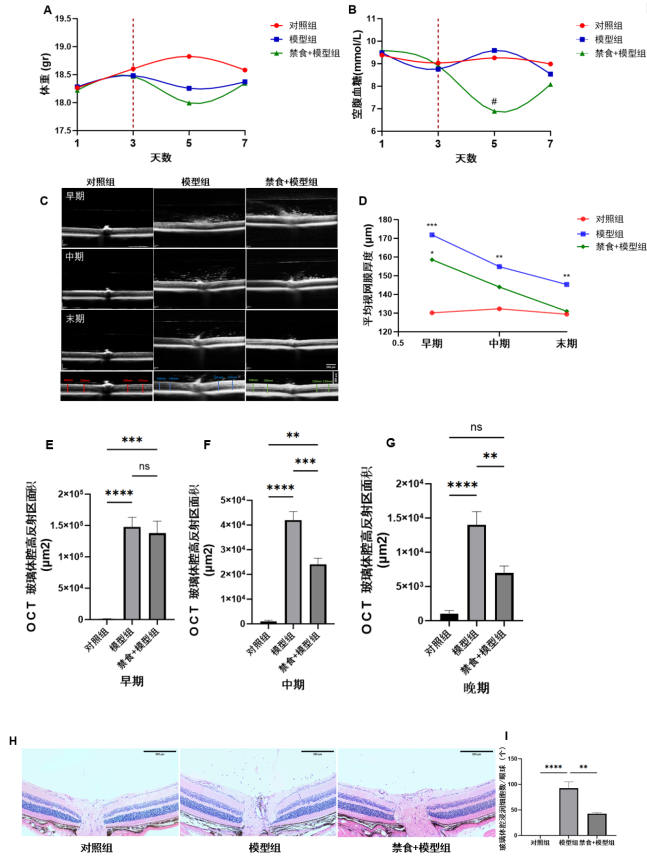
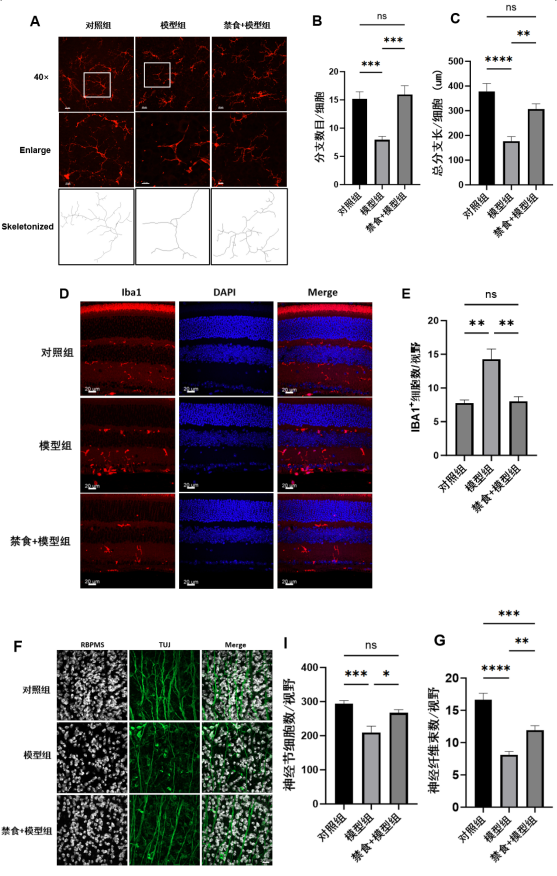
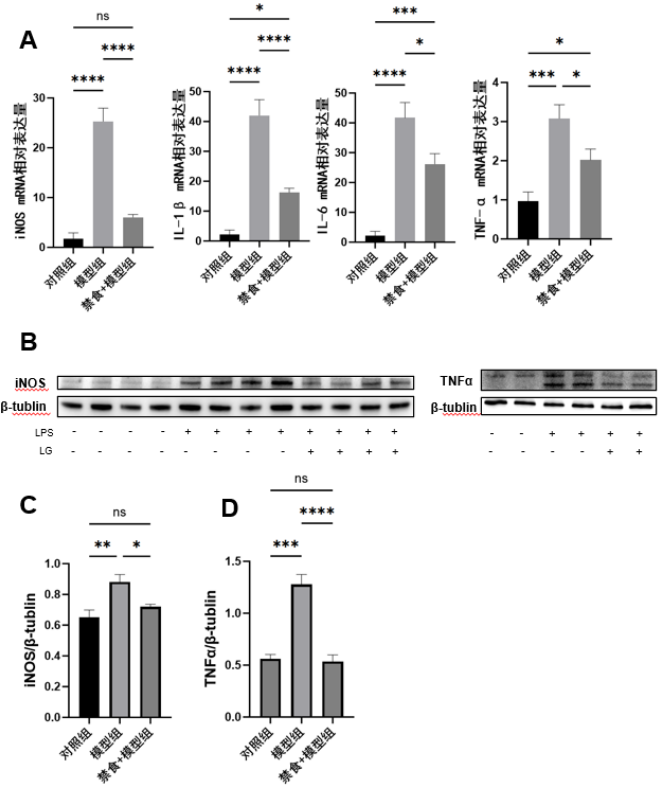
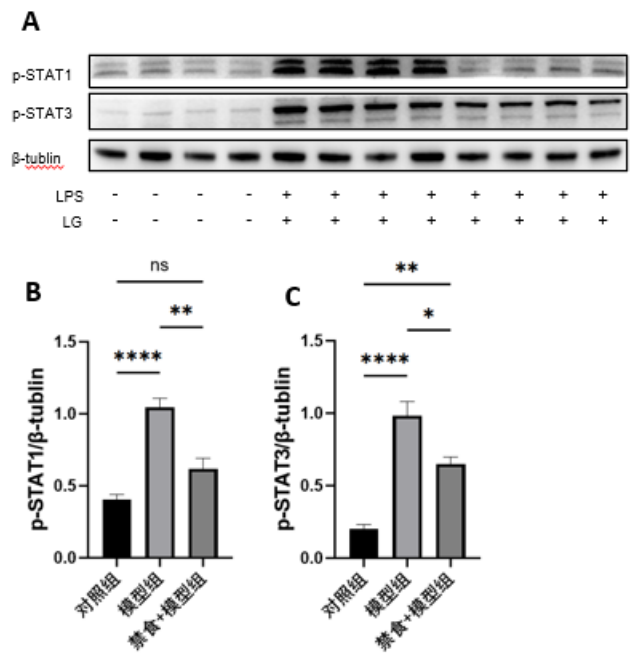
1、Krishna U, Ajanaku D, Denniston AK, et al. Uveitis: a sight-threatening
disease which can impact all systems[ J].Postgrad Med J, 2017,
93(1106): 766-773. DOI: 10.1136/postgradmedj-2017-134891.Krishna U, Ajanaku D, Denniston AK, et al. Uveitis: a sight-threatening
disease which can impact all systems[ J].Postgrad Med J, 2017,
93(1106): 766-773. DOI: 10.1136/postgradmedj-2017-134891.
2、Ross BX, Habhab S, Syeda S, et al. Patient clinical outcomes in
standalone versus a combined ophthalmology-rheumatology uveitis
clinic[ J]. J Ophthal Inflamm Infect, 2022, 12: 36. DOI: 10.1186/
s12348-022-00314-1.Ross BX, Habhab S, Syeda S, et al. Patient clinical outcomes in
standalone versus a combined ophthalmology-rheumatology uveitis
clinic[ J]. J Ophthal Inflamm Infect, 2022, 12: 36. DOI: 10.1186/
s12348-022-00314-1.
3、Foster CS, Kothari S, Anesi SD, et al. The Ocular Immunology
and Uveitis Foundation preferred practice patterns of uveitis
management[ J].Surv Ophthalmol, 2016, 61(1): 1-17. DOI: 10.1016/
j.survophthal.2015.07.001.Foster CS, Kothari S, Anesi SD, et al. The Ocular Immunology
and Uveitis Foundation preferred practice patterns of uveitis
management[ J].Surv Ophthalmol, 2016, 61(1): 1-17. DOI: 10.1016/
j.survophthal.2015.07.001.
4、Mi l l s SA , Jobling AI, Di xonM A , et al. Fractal k ine-induced
microglial vasoregulation occurs within the retina and is altered
early in diabetic retinopathy[ J]. Proc Natl Acad Sci U S A, 2021,
118(51):e2112561118.. DOI: 10.1073/pnas.2112561118.Mi l l s SA , Jobling AI, Di xonM A , et al. Fractal k ine-induced
microglial vasoregulation occurs within the retina and is altered
early in diabetic retinopathy[ J]. Proc Natl Acad Sci U S A, 2021,
118(51):e2112561118.. DOI: 10.1073/pnas.2112561118.
5、Okunuki Y, Mukai R , NakaoT, et al. Retinal microglia initiate
neuroinflammation in ocular autoimmunity[ J]. Proc Natl Acad Sci U S
A, 2019, 116(20): 9989-9998. DOI: 10.1073/pnas.1820387116.Okunuki Y, Mukai R , NakaoT, et al. Retinal microglia initiate
neuroinflammation in ocular autoimmunity[ J]. Proc Natl Acad Sci U S
A, 2019, 116(20): 9989-9998. DOI: 10.1073/pnas.1820387116.
6、Kokona D, Ebneter A, Escher P, et al. Colony-stimulating factor 1
receptor inhibition prevents disruption of the blood-retina barrier
during chronic inflammation[ J].J Neuroinflammation, 2018, 15(1):
340. DOI: 10.1186/s12974-018-1373-4.Kokona D, Ebneter A, Escher P, et al. Colony-stimulating factor 1
receptor inhibition prevents disruption of the blood-retina barrier
during chronic inflammation[ J].J Neuroinflammation, 2018, 15(1):
340. DOI: 10.1186/s12974-018-1373-4.
7、Conrady CD, Yeh S. A review of ocular drug delivery platforms and
drugs for infectious and noninfectious uveitis: the past, present,
and future[ J]. Pharmaceutics, 2021, 13(8):1224. DOI: 10.3390/
pharmaceutics13081224.Conrady CD, Yeh S. A review of ocular drug delivery platforms and
drugs for infectious and noninfectious uveitis: the past, present,
and future[ J]. Pharmaceutics, 2021, 13(8):1224. DOI: 10.3390/
pharmaceutics13081224.
8、Wu X, TaoM, ZhuL, et al. Pathogenesis and current therapies for non�infectious uveitis[ J].Clin Exp Med, 2023, 23(4): 1089-1106. DOI:
10.1007/s10238-022-00954-6.Wu X, TaoM, ZhuL, et al. Pathogenesis and current therapies for non�infectious uveitis[ J].Clin Exp Med, 2023, 23(4): 1089-1106. DOI:
10.1007/s10238-022-00954-6.
9、Cignarella F, Cantoni C, Ghezzi L, et al. Intermittent fasting confers
protection in CNS autoimmunity by altering the gut microbiota[ J].Cell
Metab, 2018, 27(6): 1222-1235.e6. DOI: 10.1016/j.cmet.2018.05.006.Cignarella F, Cantoni C, Ghezzi L, et al. Intermittent fasting confers
protection in CNS autoimmunity by altering the gut microbiota[ J].Cell
Metab, 2018, 27(6): 1222-1235.e6. DOI: 10.1016/j.cmet.2018.05.006.
10、Faris AIE, Kacimi S, Al-Kurd RA, et al. Intermittent fasting during
Ramadan attenuates proinflammatory cytokines and immune cells in
healthy subjects[ J].Nutr Res, 2012, 32(12): 947-955. DOI: 10.1016/
j.nutres.2012.06.021.Faris AIE, Kacimi S, Al-Kurd RA, et al. Intermittent fasting during
Ramadan attenuates proinflammatory cytokines and immune cells in
healthy subjects[ J].Nutr Res, 2012, 32(12): 947-955. DOI: 10.1016/
j.nutres.2012.06.021.
11、Dong%20TA%2C%20Sandesara%20PB%2C%20Dhindsa%20DS%2C%20et%20al.%20Intermittent%20fasting%3A%20aheart%20%0Ahealthy%20dietary%20pattern%3F%5B%20J%5D.Am%20J%20Med%2C%202020%2C%20133(8)%3A%20901-907.%20DOI%3A%20%0A10.1016%2Fj.amjmed.2020.03.030.Dong%20TA%2C%20Sandesara%20PB%2C%20Dhindsa%20DS%2C%20et%20al.%20Intermittent%20fasting%3A%20aheart%20%0Ahealthy%20dietary%20pattern%3F%5B%20J%5D.Am%20J%20Med%2C%202020%2C%20133(8)%3A%20901-907.%20DOI%3A%20%0A10.1016%2Fj.amjmed.2020.03.030.
12、Han X, Xu T, Fang Q, et al. Quercetin hinders microglial activation to
alleviate neurotoxicity via the interplay between NLRP3 inflammasome
and mitophagy[ J].Redox Biol,2021, 44 : 102010.DOI: 10.1016/
j.redox.2021.102010.Han X, Xu T, Fang Q, et al. Quercetin hinders microglial activation to
alleviate neurotoxicity via the interplay between NLRP3 inflammasome
and mitophagy[ J].Redox Biol,2021, 44 : 102010.DOI: 10.1016/
j.redox.2021.102010.
13、Koga T, Koshiyama Y, Gotoh T, et al. Coinduction of nitric oxide
synthase and arginine metabolic enzymes in endotoxin-induced
uveitis rats[ J].Exp Eye Res, 2002, 75(6): 659-667. DOI: 10.1006/
exer.2002.2062.Koga T, Koshiyama Y, Gotoh T, et al. Coinduction of nitric oxide
synthase and arginine metabolic enzymes in endotoxin-induced
uveitis rats[ J].Exp Eye Res, 2002, 75(6): 659-667. DOI: 10.1006/
exer.2002.2062.
14、Egwuagu CE, Alhakeem SA , Mbanefo EC. Uveitis: molecular
pathogenesis and emerging therapies[ J]. Front Immunol, 2021,
12:623725. DOI:10.3389/fimmu.2021.623725.Egwuagu CE, Alhakeem SA , Mbanefo EC. Uveitis: molecular
pathogenesis and emerging therapies[ J]. Front Immunol, 2021,
12:623725. DOI:10.3389/fimmu.2021.623725.
15、Hatori M, Vollmers C, Zarrinpar A, et al. Time-restricted feeding
without reducing caloric intake prevents metabolic diseases in mice fed
a high-fat diet[ J].Cell Metab, 2012, 15(6): 848-860. DOI: 10.1016/
j.cmet.2012.04.019.Hatori M, Vollmers C, Zarrinpar A, et al. Time-restricted feeding
without reducing caloric intake prevents metabolic diseases in mice fed
a high-fat diet[ J].Cell Metab, 2012, 15(6): 848-860. DOI: 10.1016/
j.cmet.2012.04.019.
16、Mager DE, WanR, BrownM, et al. Caloric restriction and intermittent
fasting alter spectral measures of heart rate and blood pressure
variability in rats[ J]. FASEB J, 2006, 20(6): 631-637. DOI: 10.1096/
fj.05-5263com.Mager DE, WanR, BrownM, et al. Caloric restriction and intermittent
fasting alter spectral measures of heart rate and blood pressure
variability in rats[ J]. FASEB J, 2006, 20(6): 631-637. DOI: 10.1096/
fj.05-5263com.
17、Vasim I, Majeed CN, DeBoer MD. Intermittent fasting and metabolic
health[ J]. Nutrients, 2022, 14(3):631. DOI: 10.3390/nu14030631.Vasim I, Majeed CN, DeBoer MD. Intermittent fasting and metabolic
health[ J]. Nutrients, 2022, 14(3):631. DOI: 10.3390/nu14030631.
18、Liu S, Zeng M, WanW, et al. The health-promoting effects and the
mechanism of intermittent fasting[ J].J Diabetes Res, 2023, 2023:
4038546. DOI:10.1155/2023/4038546.Liu S, Zeng M, WanW, et al. The health-promoting effects and the
mechanism of intermittent fasting[ J].J Diabetes Res, 2023, 2023:
4038546. DOI:10.1155/2023/4038546.
19、Wang L, WangYJ, WangR, et al. Fasting mimicking diet inhibits tumor�associated macrophage survival and pro-tumor function in hypoxia:
implications for combination therapy with anti-angiogenic agent[ J].J
Transl Med, 2023, 21(1): 754. DOI: 10.1186/s12967-023-04577-7.Wang L, WangYJ, WangR, et al. Fasting mimicking diet inhibits tumor�associated macrophage survival and pro-tumor function in hypoxia:
implications for combination therapy with anti-angiogenic agent[ J].J
Transl Med, 2023, 21(1): 754. DOI: 10.1186/s12967-023-04577-7.
20、Jordan S, Tung N, Casanova-Acebes M, et al. Dietary intake regulates
the circulating inflammatory monocyte pool[ J]. Cell, 2019, 178(5):
1102-1114.e17. DOI: 10.1016/j.cell.2019.07.050.Jordan S, Tung N, Casanova-Acebes M, et al. Dietary intake regulates
the circulating inflammatory monocyte pool[ J]. Cell, 2019, 178(5):
1102-1114.e17. DOI: 10.1016/j.cell.2019.07.050.
21、Orihuela R, McPherson CA, Harry GJ. Microglial M1/M2 polarization
and metabolic states[ J].British J Pharmacology, 2016, 173(4): 649-
665. DOI: 10.1111/bph.13139.Orihuela R, McPherson CA, Harry GJ. Microglial M1/M2 polarization
and metabolic states[ J].British J Pharmacology, 2016, 173(4): 649-
665. DOI: 10.1111/bph.13139.
22、Song MY, Yi F, Xiao H, et al.Energy restriction induced SIRT6
inhibits microglia activation and promotes angiogenesis in cerebral
ischemia via transcriptional inhibition of TXNIP[ J]. Cell Death
Dis,2022,13(5):449.DOI: 10.1038/s41419-022-04866-x.Song MY, Yi F, Xiao H, et al.Energy restriction induced SIRT6
inhibits microglia activation and promotes angiogenesis in cerebral
ischemia via transcriptional inhibition of TXNIP[ J]. Cell Death
Dis,2022,13(5):449.DOI: 10.1038/s41419-022-04866-x.
23、Kim KY, Ju WK, Neufeld AH. Neuronal susceptibility to damage:
comparison of the retinas of young, old and old/caloric restricted rats
before and after transient ischemia[ J].Neurobiol Aging, 2004, 25(4):
491-500. DOI: 10.1016/j.neurobiolaging.2003.07.005.Kim KY, Ju WK, Neufeld AH. Neuronal susceptibility to damage:
comparison of the retinas of young, old and old/caloric restricted rats
before and after transient ischemia[ J].Neurobiol Aging, 2004, 25(4):
491-500. DOI: 10.1016/j.neurobiolaging.2003.07.005.
24、Zarrinpar A, Chaix A, Yooseph S, et al. Diet and feeding pattern affect
the diurnal dynamics of the gut microbiome[ J].Cell Metab, 2014,
20(6): 1006-1017. DOI: 10.1016/j.cmet.2014.11.008.Zarrinpar A, Chaix A, Yooseph S, et al. Diet and feeding pattern affect
the diurnal dynamics of the gut microbiome[ J].Cell Metab, 2014,
20(6): 1006-1017. DOI: 10.1016/j.cmet.2014.11.008.
25、Galdiero M, Vitiello M, D’Isanto M, et al. STAT1 and STAT3
phosphorylation by porins are independent of JAKs but are dependent
on MAPK pathway and plays a role in U937 cells production of
interleukin-6[ J]. Cytokine, 2006, 36(5-6): 218-228. DOI: 10.1016/
j.cyto.2006.12.003.Galdiero M, Vitiello M, D’Isanto M, et al. STAT1 and STAT3
phosphorylation by porins are independent of JAKs but are dependent
on MAPK pathway and plays a role in U937 cells production of
interleukin-6[ J]. Cytokine, 2006, 36(5-6): 218-228. DOI: 10.1016/
j.cyto.2006.12.003.
26、Liang YB, TangH, ChenZB, et al. Downregulated SOCS1 expression
activates the JAK1/STAT1 pathway and promotes polarization of
macrophages into M1 type[ J].Mol Med Rep, 2017, 16(5): 6405-6411.
DOI: 10.3892/mmr.2017.7384.Liang YB, TangH, ChenZB, et al. Downregulated SOCS1 expression
activates the JAK1/STAT1 pathway and promotes polarization of
macrophages into M1 type[ J].Mol Med Rep, 2017, 16(5): 6405-6411.
DOI: 10.3892/mmr.2017.7384.
27、DongYF, NiuCS, Wang Y, et al. SOCS1/JAK2/STAT3 axis regulates
early brain injur y induced by subarachnoid hemorrhage v ia
inflammatory responses[ J]. Neural Regen Res, 2021, 16(12): 2453.
DOI: 10.4103/1673-5374.313049.DongYF, NiuCS, Wang Y, et al. SOCS1/JAK2/STAT3 axis regulates
early brain injur y induced by subarachnoid hemorrhage v ia
inflammatory responses[ J]. Neural Regen Res, 2021, 16(12): 2453.
DOI: 10.4103/1673-5374.313049.
28、Feng X, Li M, Lin Z, et al. Tetramethylpyrazine promotes axonal
remodeling and modulates microglial polarization via JAK2-STAT1/3
and GSK3-NFkappaB pathways in ischemic stroke, Neurochem
Int[ J].2023,170:105607.DOI: 10.1016/j.neuint.2023.105607.Feng X, Li M, Lin Z, et al. Tetramethylpyrazine promotes axonal
remodeling and modulates microglial polarization via JAK2-STAT1/3
and GSK3-NFkappaB pathways in ischemic stroke, Neurochem
Int[ J].2023,170:105607.DOI: 10.1016/j.neuint.2023.105607.
29、Ma Y, TangT, ShengL, et al. Aloin suppresses lipopolysaccharide�induced inflammation by inhibiting JAK1-STAT1/3 activation
and ROS production in R AW264.7 cells[ J]. Int J Mol Med,
2018,42(4):1925-193. DOI: 10.3892/ijmm.2018.3796.Ma Y, TangT, ShengL, et al. Aloin suppresses lipopolysaccharide�induced inflammation by inhibiting JAK1-STAT1/3 activation
and ROS production in R AW264.7 cells[ J]. Int J Mol Med,
2018,42(4):1925-193. DOI: 10.3892/ijmm.2018.3796.