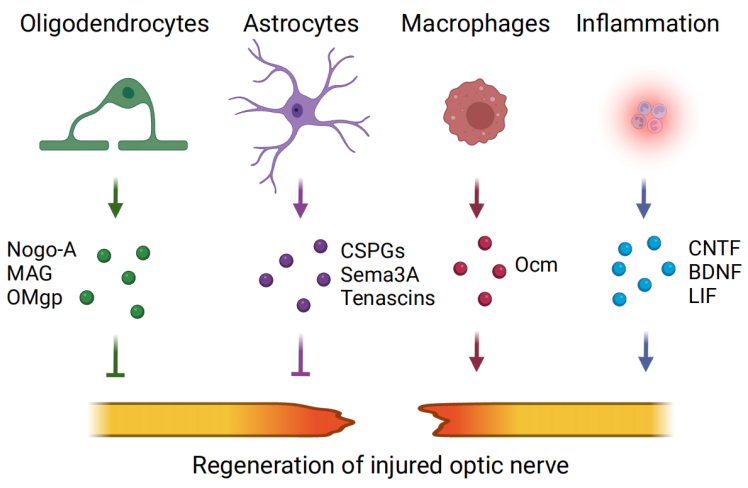
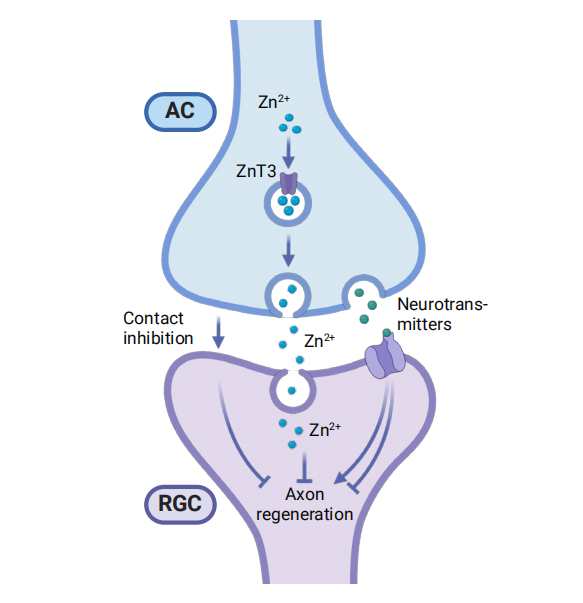
1、Dhande OS, Stafford BK, Lim JH A, et al. Contributions
of retinal ganglion cells to subcortical visual processing
and behaviors. Annu Rev Vis Sci. 2015, 1: 291-328. DOI:
10.1146/annurev-vision-082114-035502.Dhande OS, Stafford BK, Lim JH A, et al. Contributions
of retinal ganglion cells to subcortical visual processing
and behaviors. Annu Rev Vis Sci. 2015, 1: 291-328. DOI:
10.1146/annurev-vision-082114-035502.
2、Moore DL, Goldberg JL. Four steps to optic nerve
regeneration. J Neuroophthalmol. 2010, 30(4): 347-360.
DOI: 10.1097/WNO.0b013e3181e755af.Moore DL, Goldberg JL. Four steps to optic nerve
regeneration. J Neuroophthalmol. 2010, 30(4): 347-360.
DOI: 10.1097/WNO.0b013e3181e755af.
3、Laha B, Stafford BK, Huberman AD. Regenerating
optic pathways from the eye to the brain. Science. 2017,
356(6342): 1031-1034. DOI: 10.1126/science.aal5060.Laha B, Stafford BK, Huberman AD. Regenerating
optic pathways from the eye to the brain. Science. 2017,
356(6342): 1031-1034. DOI: 10.1126/science.aal5060.
4、Popovich%20PG%2C%20Longbrake%20EE.%20Can%20the%20immune%20system%20be%20%0Aharnessed%20to%20repair%20the%20CNS%3F.%20Nat%20Rev%20Neurosci.%202008%2C%20%0A9(6)%3A%20481-493.%20DOI%3A%2010.1038%2Fnrn2398.Popovich%20PG%2C%20Longbrake%20EE.%20Can%20the%20immune%20system%20be%20%0Aharnessed%20to%20repair%20the%20CNS%3F.%20Nat%20Rev%20Neurosci.%202008%2C%20%0A9(6)%3A%20481-493.%20DOI%3A%2010.1038%2Fnrn2398.
5、Yiu G, He Z. Glial inhibition of CNS axon regeneration.
Nat Rev Neurosci. 2006, 7(8): 617-627. DOI: 10.1038/
nrn1956.Yiu G, He Z. Glial inhibition of CNS axon regeneration.
Nat Rev Neurosci. 2006, 7(8): 617-627. DOI: 10.1038/
nrn1956.
6、Kurimoto T, Yin Y, Habboub G, et al. Neutrophils express
oncomodulin and promote optic nerve regeneration. J
Neurosci. 2013, 33(37): 14816-14824. DOI: 10.1523/
JNEUROSCI.5511-12.2013.Kurimoto T, Yin Y, Habboub G, et al. Neutrophils express
oncomodulin and promote optic nerve regeneration. J
Neurosci. 2013, 33(37): 14816-14824. DOI: 10.1523/
JNEUROSCI.5511-12.2013.
7、Sergeeva EG, Rosenberg PA, Benowitz LI. Non-cell�autonomous regulation of optic nerve regeneration by
amacrine cells. Front Cell Neurosci. 2021, 15: 666798.
DOI: 10.3389/fncel.2021.666798.Sergeeva EG, Rosenberg PA, Benowitz LI. Non-cell�autonomous regulation of optic nerve regeneration by
amacrine cells. Front Cell Neurosci. 2021, 15: 666798.
DOI: 10.3389/fncel.2021.666798.
8、Lee Y, Morrison BM, Li Y, et al. Oligodendroglia
metabolically support axons and contribute to
neurodegeneration. Nature. 2012, 487(7408): 443-448.
DOI: 10.1038/nature11314.Lee Y, Morrison BM, Li Y, et al. Oligodendroglia
metabolically support axons and contribute to
neurodegeneration. Nature. 2012, 487(7408): 443-448.
DOI: 10.1038/nature11314.
9、Chen MS, Huber AB, van der Haar ME, et al. Nogo-a
is a myelin-associated neurite outgrowth inhibitor and
an antigen for monoclonal antibody IN-1. Nature. 2000,
403(6768): 434-439. DOI: 10.1038/35000219.Chen MS, Huber AB, van der Haar ME, et al. Nogo-a
is a myelin-associated neurite outgrowth inhibitor and
an antigen for monoclonal antibody IN-1. Nature. 2000,
403(6768): 434-439. DOI: 10.1038/35000219.
10、GrandPré T, Li S, Strittmatter SM. Nogo-66 receptor
antagonist peptide promotes axonal regeneration. Nature.
2002, 417(6888): 547-551. DOI: 10.1038/417547a.GrandPré T, Li S, Strittmatter SM. Nogo-66 receptor
antagonist peptide promotes axonal regeneration. Nature.
2002, 417(6888): 547-551. DOI: 10.1038/417547a.
11、Wang KC, Koprivica V, Kim JA, et al. Oligodendrocyte�myelin glycoprotein is a Nogo receptor ligand that inhibits
neurite outgrowth. Nature. 2002, 417(6892): 941-944.
DOI: 10.1038/nature00867.Wang KC, Koprivica V, Kim JA, et al. Oligodendrocyte�myelin glycoprotein is a Nogo receptor ligand that inhibits
neurite outgrowth. Nature. 2002, 417(6892): 941-944.
DOI: 10.1038/nature00867.
12、Kottis V, Thibault P, Mikol D, et al. Oligodendrocyte�myelin glycoprotein (OMgp) is an inhibitor of neurite
outgrowth. J Neurochem. 2002, 82(6): 1566-1569. DOI:
10.1046/j.1471-4159.2002.01146.x.Kottis V, Thibault P, Mikol D, et al. Oligodendrocyte�myelin glycoprotein (OMgp) is an inhibitor of neurite
outgrowth. J Neurochem. 2002, 82(6): 1566-1569. DOI:
10.1046/j.1471-4159.2002.01146.x.
13、Atwal JK, Pinkston-Gosse J, Syken J, et al. PirB is
a functional receptor for myelin inhibitors of axonal
regeneration. Science. 2008, 322(5903): 967-970. DOI:
10.1126/science.1161151.Atwal JK, Pinkston-Gosse J, Syken J, et al. PirB is
a functional receptor for myelin inhibitors of axonal
regeneration. Science. 2008, 322(5903): 967-970. DOI:
10.1126/science.1161151.
14、Filbin MT. PirB, a second receptor for the myelin
inhibitors of axonal regeneration Nogo66, MAG, and
OMgp: implications for regeneration in vivo. Neuron.
2008, 60(5): 740-742. DOI: 10.1016/j.neuron.2008.12.001.Filbin MT. PirB, a second receptor for the myelin
inhibitors of axonal regeneration Nogo66, MAG, and
OMgp: implications for regeneration in vivo. Neuron.
2008, 60(5): 740-742. DOI: 10.1016/j.neuron.2008.12.001.
15、Domeniconi M, Cao Z, Spencer T, et al. Myelin-associated
glycoprotein interacts with the Nogo66 receptor to inhibit
neurite outgrowth. Neuron. 2002, 35(2): 283-290. DOI:
10.1016/s0896-6273(02)00770-5.Domeniconi M, Cao Z, Spencer T, et al. Myelin-associated
glycoprotein interacts with the Nogo66 receptor to inhibit
neurite outgrowth. Neuron. 2002, 35(2): 283-290. DOI:
10.1016/s0896-6273(02)00770-5.
16、Liu BP, Fournier A, GrandPré T, et al. Myelin-associated
glycoprotein as a functional ligand for the Nogo-66
receptor. Science. 2002, 297(5584): 1190-1193. DOI:
10.1126/science.1073031.Liu BP, Fournier A, GrandPré T, et al. Myelin-associated
glycoprotein as a functional ligand for the Nogo-66
receptor. Science. 2002, 297(5584): 1190-1193. DOI:
10.1126/science.1073031.
17、Mi S, Lee X, Shao Z, et al. LINGO-1 is a component
of the Nogo-66 receptor/p75 signaling complex. Nat
Neurosci. 2004, 7(3): 221-228. DOI: 10.1038/nn1188.Mi S, Lee X, Shao Z, et al. LINGO-1 is a component
of the Nogo-66 receptor/p75 signaling complex. Nat
Neurosci. 2004, 7(3): 221-228. DOI: 10.1038/nn1188.
18、Wong ST, Henley JR, Kanning KC, et al. A p75(NTR)
and Nogo receptor complex mediates repulsive signaling
by myelin-associated glycoprotein. Nat Neurosci. 2002,
5(12): 1302-1308. DOI: 10.1038/nn975.Wong ST, Henley JR, Kanning KC, et al. A p75(NTR)
and Nogo receptor complex mediates repulsive signaling
by myelin-associated glycoprotein. Nat Neurosci. 2002,
5(12): 1302-1308. DOI: 10.1038/nn975.
19、Venkatesh K, Chivatakarn O, Lee H, et al. The Nogo-
66 receptor homolog NgR2 is a sialic acid-dependent
receptor selective for myelin-associated glycoprotein.
J Neurosci. 2005, 25(4): 808-822. DOI: 10.1523/JNEUROSCI.4464-04.2005.Venkatesh K, Chivatakarn O, Lee H, et al. The Nogo-
66 receptor homolog NgR2 is a sialic acid-dependent
receptor selective for myelin-associated glycoprotein.
J Neurosci. 2005, 25(4): 808-822. DOI: 10.1523/JNEUROSCI.4464-04.2005.
20、Lee JK, Geoffroy CG, Chan AF, et al. Assessing spinal
axon regeneration and sprouting in Nogo-, MAG-, and
OMgp-deficient mice. Neuron. 2010, 66(5): 663-670. DOI:
10.1016/j.neuron.2010.05.002.Lee JK, Geoffroy CG, Chan AF, et al. Assessing spinal
axon regeneration and sprouting in Nogo-, MAG-, and
OMgp-deficient mice. Neuron. 2010, 66(5): 663-670. DOI:
10.1016/j.neuron.2010.05.002.
21、Cafferty WBJ, Duffy P, Huebner E, et al. MAG and
OMgp synergize with Nogo-A to restrict axonal growth
and neurological recovery after spinal cord trauma. J
Neurosci. 2010, 30(20): 6825-6837. DOI: 10.1523/
JNEUROSCI.6239-09.2010.Cafferty WBJ, Duffy P, Huebner E, et al. MAG and
OMgp synergize with Nogo-A to restrict axonal growth
and neurological recovery after spinal cord trauma. J
Neurosci. 2010, 30(20): 6825-6837. DOI: 10.1523/
JNEUROSCI.6239-09.2010.
22、Vajda F, Jordi N, Dalkara D, et al. Cell type-specific
Nogo-a gene ablation promotes axonal regeneration in the
injured adult optic nerve. Cell Death Differ. 2015, 22(2):
323-335. DOI: 10.1038/cdd.2014.147.Vajda F, Jordi N, Dalkara D, et al. Cell type-specific
Nogo-a gene ablation promotes axonal regeneration in the
injured adult optic nerve. Cell Death Differ. 2015, 22(2):
323-335. DOI: 10.1038/cdd.2014.147.
23、Liebscher T, Schnell L, Schnell D, et al. Nogo-a antibody
improves regeneration and locomotion of spinal cord�injured rats. Ann Neurol. 2005, 58(5): 706-719. DOI:
10.1002/ana.20627.Liebscher T, Schnell L, Schnell D, et al. Nogo-a antibody
improves regeneration and locomotion of spinal cord�injured rats. Ann Neurol. 2005, 58(5): 706-719. DOI:
10.1002/ana.20627.
24、Merkler D, Metz GA, Raineteau O, et al. Locomotor
recovery in spinal cord-injured rats treated with an
antibody neutralizing the myelin-associated neurite growth
inhibitor Nogo-A. J Neurosci. 2001, 21(10): 3665-3673.
DOI: 10.1523/JNEUROSCI.21-10-03665.2001.Merkler D, Metz GA, Raineteau O, et al. Locomotor
recovery in spinal cord-injured rats treated with an
antibody neutralizing the myelin-associated neurite growth
inhibitor Nogo-A. J Neurosci. 2001, 21(10): 3665-3673.
DOI: 10.1523/JNEUROSCI.21-10-03665.2001.
25、Zheng B, Atwal J, Ho C, et al. Genetic deletion of the
Nogo receptor does not reduce neurite inhibition in vitro or
promote corticospinal tract regeneration in vivo. Proc Natl
Acad Sci U S A. 2005, 102(4): 1205-1210. DOI: 10.1073/
pnas.0409026102.Zheng B, Atwal J, Ho C, et al. Genetic deletion of the
Nogo receptor does not reduce neurite inhibition in vitro or
promote corticospinal tract regeneration in vivo. Proc Natl
Acad Sci U S A. 2005, 102(4): 1205-1210. DOI: 10.1073/
pnas.0409026102.
26、Dickendesher TL, Baldwin KT, Mironova YA, et al.
NgR1 and NgR3 are receptors for chondroitin sulfate
proteoglycans. Nat Neurosci. 2012, 15(5): 703-712. DOI:
10.1038/nn.3070.Dickendesher TL, Baldwin KT, Mironova YA, et al.
NgR1 and NgR3 are receptors for chondroitin sulfate
proteoglycans. Nat Neurosci. 2012, 15(5): 703-712. DOI:
10.1038/nn.3070.
27、Kawakami Y, Kurihara Y, Saito Y, et al. The soluble form
of LOTUS inhibits nogo receptor-mediated signaling by
interfering with the interaction between nogo receptor type
1 and p75 neurotrophin receptor. J Neurosci. 2018, 38(10):2589-2604. DOI: 10.1523/JNEUROSCI.0953-17.2018.Kawakami Y, Kurihara Y, Saito Y, et al. The soluble form
of LOTUS inhibits nogo receptor-mediated signaling by
interfering with the interaction between nogo receptor type
1 and p75 neurotrophin receptor. J Neurosci. 2018, 38(10):2589-2604. DOI: 10.1523/JNEUROSCI.0953-17.2018.
28、Kurihara Y, Takai T, Takei K. Nogo receptor antagonist
LOTUS exerts suppression on axonal growth-inhibiting
receptor PIR-B. J Neurochem. 2020, 155(3): 285-299.
DOI: 10.1111/jnc.15013.Kurihara Y, Takai T, Takei K. Nogo receptor antagonist
LOTUS exerts suppression on axonal growth-inhibiting
receptor PIR-B. J Neurochem. 2020, 155(3): 285-299.
DOI: 10.1111/jnc.15013.
29、Su Y, Yu Z, Guo X, et al. Axonal regeneration of optic
nerve after crush after PirBsiRNA transfection. Int J Clin
Exp Pathol. 2017, 10(9): 9633-9638.Su Y, Yu Z, Guo X, et al. Axonal regeneration of optic
nerve after crush after PirBsiRNA transfection. Int J Clin
Exp Pathol. 2017, 10(9): 9633-9638.
30、Yang M, Jian L, Fan W, et al. Axon regeneration after optic
nerve injury in rats can be improved via PirB knockdown
in the retina. Cell Biosci. 2021, 11(1): 158. DOI: 10.1186/
s13578-021-00670-w.Yang M, Jian L, Fan W, et al. Axon regeneration after optic
nerve injury in rats can be improved via PirB knockdown
in the retina. Cell Biosci. 2021, 11(1): 158. DOI: 10.1186/
s13578-021-00670-w.
31、Sofroniew MV. Molecular dissection of reactive
astrogliosis and glial scar formation. Trends Neurosci.
2009, 32(12): 638-647. DOI: 10.1016/j.tins.2009.08.002.Sofroniew MV. Molecular dissection of reactive
astrogliosis and glial scar formation. Trends Neurosci.
2009, 32(12): 638-647. DOI: 10.1016/j.tins.2009.08.002.
32、Carulli D, Rhodes KE, Brown DJ, et al. Composition
of perineuronal nets in the adult rat cerebellum and the
cellular origin of their components. J Comp Neurol. 2006,
494(4): 559-577. DOI: 10.1002/cne.20822.Carulli D, Rhodes KE, Brown DJ, et al. Composition
of perineuronal nets in the adult rat cerebellum and the
cellular origin of their components. J Comp Neurol. 2006,
494(4): 559-577. DOI: 10.1002/cne.20822.
33、De Winter F, Oudega M, Lankhorst AJ, et al. Injury�induced class 3 semaphorin expression in the rat spinal
cord. Exp Neurol. 2002, 175(1): 61-75. DOI: 10.1006/
exnr.2002.7884.De Winter F, Oudega M, Lankhorst AJ, et al. Injury�induced class 3 semaphorin expression in the rat spinal
cord. Exp Neurol. 2002, 175(1): 61-75. DOI: 10.1006/
exnr.2002.7884.
34、Kaneko S, Iwanami A, Nakamura M, et al. A selective
Sema3A inhibitor enhances regenerative responses and
functional recovery of the injured spinal cord. Nat Med.
2006, 12(12): 1380-1389. DOI: 10.1038/nm1505.Kaneko S, Iwanami A, Nakamura M, et al. A selective
Sema3A inhibitor enhances regenerative responses and
functional recovery of the injured spinal cord. Nat Med.
2006, 12(12): 1380-1389. DOI: 10.1038/nm1505.
35、Apostolova I, Irintchev A, Schachner M. Tenascin-R
restricts posttraumatic remodeling of motoneuron
innervation and functional recovery after spinal cord injury
in adult mice. J Neurosci. 2006, 26(30): 7849-7859. DOI:
10.1523/JNEUROSCI.1526-06.2006.Apostolova I, Irintchev A, Schachner M. Tenascin-R
restricts posttraumatic remodeling of motoneuron
innervation and functional recovery after spinal cord injury
in adult mice. J Neurosci. 2006, 26(30): 7849-7859. DOI:
10.1523/JNEUROSCI.1526-06.2006.
36、Shen Y, Tenney AP, Busch SA, et al. PTPsigma is a
receptor for chondroitin sulfate proteoglycan, an inhibitor
of neural regeneration. Science. 2009, 326(5952): 592-
596. DOI: 10.1126/science.1178310.Shen Y, Tenney AP, Busch SA, et al. PTPsigma is a
receptor for chondroitin sulfate proteoglycan, an inhibitor
of neural regeneration. Science. 2009, 326(5952): 592-
596. DOI: 10.1126/science.1178310.
37、Torres-Vázquez J, Gitler AD, Fraser SD, et al. Semaphorin�plexin signaling guides patterning of the developing
vasculature. Dev Cell. 2004, 7(1): 117-123. DOI: 10.1016/
j.devcel.2004.06.008.Torres-Vázquez J, Gitler AD, Fraser SD, et al. Semaphorin�plexin signaling guides patterning of the developing
vasculature. Dev Cell. 2004, 7(1): 117-123. DOI: 10.1016/
j.devcel.2004.06.008.
38、Andrews MR, Czvitkovich S, Dassie E, et al.
Alpha9 integrin promotes neurite outgrowth on
tenascin-C and enhances sensory axon regeneration.
J Neurosci. 2009, 29(17): 5546-5557. DOI: 10.1523/
JNEUROSCI.0759-09.2009.Andrews MR, Czvitkovich S, Dassie E, et al.
Alpha9 integrin promotes neurite outgrowth on
tenascin-C and enhances sensory axon regeneration.
J Neurosci. 2009, 29(17): 5546-5557. DOI: 10.1523/
JNEUROSCI.0759-09.2009.
39、Wang X, Hasan O, Arzeno A, et al. Axonal regeneration
induced by blockade of glial inhibitors coupled with
activation of intrinsic neuronal growth pathways.
Exp Neurol. 2012, 237(1): 55-69. DOI: 10.1016/
j.expneurol.2012.06.009.Wang X, Hasan O, Arzeno A, et al. Axonal regeneration
induced by blockade of glial inhibitors coupled with
activation of intrinsic neuronal growth pathways.
Exp Neurol. 2012, 237(1): 55-69. DOI: 10.1016/
j.expneurol.2012.06.009.
40、Sapieha PS, Duplan L, Uetani N, et al. Receptor protein
tyrosine phosphatase sigma inhibits axon regrowth in the
adult injured CNS. Mol Cell Neurosci. 2005, 28(4): 625-
635. DOI: 10.1016/j.mcn.2004.10.011.Sapieha PS, Duplan L, Uetani N, et al. Receptor protein
tyrosine phosphatase sigma inhibits axon regrowth in the
adult injured CNS. Mol Cell Neurosci. 2005, 28(4): 625-
635. DOI: 10.1016/j.mcn.2004.10.011.
41、 Shirvan A, Kimron M, Holdengreber V, et al. Anti�semaphorin 3A antibodies rescue retinal ganglion cells
from cell death following optic nerve axotomy. J Biol
Chem. 2002, 277(51): 49799-49807. DOI: 10.1074/jbc.
M204793200. Shirvan A, Kimron M, Holdengreber V, et al. Anti�semaphorin 3A antibodies rescue retinal ganglion cells
from cell death following optic nerve axotomy. J Biol
Chem. 2002, 277(51): 49799-49807. DOI: 10.1074/jbc.
M204793200.
42、Han F, Huo Y, Huang CJ, et al. MicroRNA-30b promotes
axon outgrowth of retinal ganglion cells by inhibiting
Semaphorin3A expression. Brain Res. 2015, 1611: 65-73.
DOI: 10.1016/j.brainres.2015.03.014.Han F, Huo Y, Huang CJ, et al. MicroRNA-30b promotes
axon outgrowth of retinal ganglion cells by inhibiting
Semaphorin3A expression. Brain Res. 2015, 1611: 65-73.
DOI: 10.1016/j.brainres.2015.03.014.
43、Schreiber J, Schachner M, Schumacher U, et al.
Extracellular matrix alterations, accelerated leukocyte
infiltration and enhanced axonal sprouting after spinal
cord hemisection in tenascin-C-deficient mice. Acta
Histochem. 2013, 115(8): 865-878. DOI: 10.1016/
j.acthis.2013.04.009.Schreiber J, Schachner M, Schumacher U, et al.
Extracellular matrix alterations, accelerated leukocyte
infiltration and enhanced axonal sprouting after spinal
cord hemisection in tenascin-C-deficient mice. Acta
Histochem. 2013, 115(8): 865-878. DOI: 10.1016/
j.acthis.2013.04.009.
44、Chen J, Joon Lee H, Jakovcevski I, et al. The extracellular
matrix glycoprotein tenascin-C is beneficial for spinal cord
regeneration. Mol Ther. 2010, 18(10): 1769-1777. DOI:10.1038/mt.2010.133.Chen J, Joon Lee H, Jakovcevski I, et al. The extracellular
matrix glycoprotein tenascin-C is beneficial for spinal cord
regeneration. Mol Ther. 2010, 18(10): 1769-1777. DOI:10.1038/mt.2010.133.
45、Cheah M, Andrews MR, Chew DJ, et al. Expression of an
activated integrin promotes long-distance sensory axon
regeneration in the spinal cord. J Neurosci. 2016, 36(27):
7283-7297. DOI: 10.1523/JNEUROSCI.0901-16.2016.Cheah M, Andrews MR, Chew DJ, et al. Expression of an
activated integrin promotes long-distance sensory axon
regeneration in the spinal cord. J Neurosci. 2016, 36(27):
7283-7297. DOI: 10.1523/JNEUROSCI.0901-16.2016.
46、Jin X, Yamashita T. Microglia in central nervous system
repair after injury. J Biochem. 2016, 159(5): 491-496.
DOI: 10.1093/jb/mvw009.Jin X, Yamashita T. Microglia in central nervous system
repair after injury. J Biochem. 2016, 159(5): 491-496.
DOI: 10.1093/jb/mvw009.
47、Prinz M, Priller J. Microglia and brain macrophages in the
molecular age: from origin to neuropsychiatric disease.
Nat Rev Neurosci. 2014, 15(5): 300-312. DOI: 10.1038/
nrn3722.Prinz M, Priller J. Microglia and brain macrophages in the
molecular age: from origin to neuropsychiatric disease.
Nat Rev Neurosci. 2014, 15(5): 300-312. DOI: 10.1038/
nrn3722.
48、Elmore MRP, Najafi AR, Koike MA, et al. Colony�stimulating factor 1 receptor signaling is necessary for
microglia viability, unmasking a microglia progenitor cell
in the adult brain. Neuron. 2014, 82(2): 380-397. DOI:
10.1016/j.neuron.2014.02.040.Elmore MRP, Najafi AR, Koike MA, et al. Colony�stimulating factor 1 receptor signaling is necessary for
microglia viability, unmasking a microglia progenitor cell
in the adult brain. Neuron. 2014, 82(2): 380-397. DOI:
10.1016/j.neuron.2014.02.040.
49、Hilla AM, Diekmann H, Fischer D. Microglia are
irrelevant for neuronal degeneration and axon regeneration
after acute injury. J Neurosci. 2017, 37(25): 6113-6124.
DOI: 10.1523/JNEUROSCI.0584-17.2017.Hilla AM, Diekmann H, Fischer D. Microglia are
irrelevant for neuronal degeneration and axon regeneration
after acute injury. J Neurosci. 2017, 37(25): 6113-6124.
DOI: 10.1523/JNEUROSCI.0584-17.2017.
50、 Leon S, Yin Y, Nguyen J, et al. Lens injury stimulates
axon regeneration in the mature rat optic nerve. J
Neurosci. 2000, 20(12): 4615-4626. DOI: 10.1523/
JNEUROSCI.20-12-04615.2000. Leon S, Yin Y, Nguyen J, et al. Lens injury stimulates
axon regeneration in the mature rat optic nerve. J
Neurosci. 2000, 20(12): 4615-4626. DOI: 10.1523/
JNEUROSCI.20-12-04615.2000.
51、Yin Y, Cui Q, Li Y, et al. Macrophage-derived
f a c t o r s s t i m u l a t e o p t i c n e r v e r e g e n e r a t i o n . J
Neurosci. 2003, 23(6): 2284-2293. DOI: 10.1523/
JNEUROSCI.23-06-02284.2003.Yin Y, Cui Q, Li Y, et al. Macrophage-derived
f a c t o r s s t i m u l a t e o p t i c n e r v e r e g e n e r a t i o n . J
Neurosci. 2003, 23(6): 2284-2293. DOI: 10.1523/
JNEUROSCI.23-06-02284.2003.
52、David S, Bouchard C, Tsatas O, et al. Macrophages can
modify the nonpermissive nature of the adult mammalian
central nervous system. Neuron. 1990, 5(4): 463-469.
DOI: 10.1016/0896-6273(90)90085-t.David S, Bouchard C, Tsatas O, et al. Macrophages can
modify the nonpermissive nature of the adult mammalian
central nervous system. Neuron. 1990, 5(4): 463-469.
DOI: 10.1016/0896-6273(90)90085-t.
53、Li Y, Irwin N, Yin Y, et al. Axon regeneration in goldfish
and rat retinal ganglion cells: differential responsiveness to
carbohydrates and cAMP. J Neurosci. 2003, 23(21): 7830-7838. DOI: 10.1523/JNEUROSCI.23-21-07830.2003.Li Y, Irwin N, Yin Y, et al. Axon regeneration in goldfish
and rat retinal ganglion cells: differential responsiveness to
carbohydrates and cAMP. J Neurosci. 2003, 23(21): 7830-7838. DOI: 10.1523/JNEUROSCI.23-21-07830.2003.
54、Yin Y, Cui Q, Gilbert HY, et al. Oncomodulin links
inflammation to optic nerve regeneration. Proc Natl Acad
Sci USA. 2009, 106(46): 19587-19592. DOI: 10.1073/
pnas.0907085106.Yin Y, Cui Q, Gilbert HY, et al. Oncomodulin links
inflammation to optic nerve regeneration. Proc Natl Acad
Sci USA. 2009, 106(46): 19587-19592. DOI: 10.1073/
pnas.0907085106.
55、Kurimoto T, Yin Y, Omura K, et al. Long-distance axon
regeneration in the mature optic nerve: contributions
of oncomodulin, cAMP, and pten gene deletion. J
Neurosci. 2010, 30(46): 15654-15663. DOI: 10.1523/
JNEUROSCI.4340-10.2010.Kurimoto T, Yin Y, Omura K, et al. Long-distance axon
regeneration in the mature optic nerve: contributions
of oncomodulin, cAMP, and pten gene deletion. J
Neurosci. 2010, 30(46): 15654-15663. DOI: 10.1523/
JNEUROSCI.4340-10.2010.
56、Xie L, Yin Y, Jayakar S, et al. The oncomodulin receptor
ArmC10 enables axon regeneration in mice after nerve
injury and neurite outgrowth in human iPSC-derived
sensory neurons. Sci Transl Med. 2023, 15(708):
eadg6241. DOI: 10.1126/scitranslmed.adg6241.Xie L, Yin Y, Jayakar S, et al. The oncomodulin receptor
ArmC10 enables axon regeneration in mice after nerve
injury and neurite outgrowth in human iPSC-derived
sensory neurons. Sci Transl Med. 2023, 15(708):
eadg6241. DOI: 10.1126/scitranslmed.adg6241.
57、B a l d w i n K T, C a r b a j a l K S , S e g a l B M , e t a l .
Neuroinflammation triggered by β-glucan/dectin-1
signaling enables CNS axon regeneration. Proc Natl
Acad Sci USA. 2015, 112(8): 2581-2586. DOI: 10.1073/
pnas.1423221112.B a l d w i n K T, C a r b a j a l K S , S e g a l B M , e t a l .
Neuroinflammation triggered by β-glucan/dectin-1
signaling enables CNS axon regeneration. Proc Natl
Acad Sci USA. 2015, 112(8): 2581-2586. DOI: 10.1073/
pnas.1423221112.
58、Xie L, Yin Y, Benowitz L. Chemokine CCL5 promotes
robust optic nerve regeneration and mediates many
of the effects of CNTF gene therapy. Proc Natl Acad
Sci USA. 2021, 118(9): e2017282118. DOI: 10.1073/
pnas.2017282118.Xie L, Yin Y, Benowitz L. Chemokine CCL5 promotes
robust optic nerve regeneration and mediates many
of the effects of CNTF gene therapy. Proc Natl Acad
Sci USA. 2021, 118(9): e2017282118. DOI: 10.1073/
pnas.2017282118.
59、Sas AR, Carbajal KS, Jerome AD, et al. A new neutrophil
subset promotes CNS neuron survival and axon
regeneration. Nat Immunol. 2020, 21(12): 1496-1505.
DOI: 10.1038/s41590-020-00813-0.Sas AR, Carbajal KS, Jerome AD, et al. A new neutrophil
subset promotes CNS neuron survival and axon
regeneration. Nat Immunol. 2020, 21(12): 1496-1505.
DOI: 10.1038/s41590-020-00813-0.
60、Jerome AD, Sas AR, Wang Y, et al. Cytokine polarized,
alternatively activated bone marrow neutrophils drive axon
regeneration. Nat Immunol. 2024, 25(6): 957-968. DOI:
10.1038/s41590-024-01836-7.Jerome AD, Sas AR, Wang Y, et al. Cytokine polarized,
alternatively activated bone marrow neutrophils drive axon
regeneration. Nat Immunol. 2024, 25(6): 957-968. DOI:
10.1038/s41590-024-01836-7.
61、Passino R, Finneran MC, Hafner H, et al. Neutrophil�inflicted vasculature damage suppresses immune-mediated
optic nerve regeneration. Cell Rep. 2024, 43(3): 113931.DOI: 10.1016/j.celrep.2024.113931.Passino R, Finneran MC, Hafner H, et al. Neutrophil�inflicted vasculature damage suppresses immune-mediated
optic nerve regeneration. Cell Rep. 2024, 43(3): 113931.DOI: 10.1016/j.celrep.2024.113931.
62、Hilla AM, Baehr A, Leibinger M, et al. CXCR4/
CXCL12-mediated entrapment of axons at the injury site
compromises optic nerve regeneration. Proc Natl Acad
Sci USA. 2021, 118(21): e2016409118. DOI: 10.1073/
pnas.2016409118.Hilla AM, Baehr A, Leibinger M, et al. CXCR4/
CXCL12-mediated entrapment of axons at the injury site
compromises optic nerve regeneration. Proc Natl Acad
Sci USA. 2021, 118(21): e2016409118. DOI: 10.1073/
pnas.2016409118.
63、Xie L, Cen LP, Li Y, et al. Monocyte-derived SDF1
supports optic nerve regeneration and alters retinal
ganglion cells' response to Pten deletion. Proc Natl Acad
Sci USA. 2022, 119(15): e2113751119. DOI: 10.1073/
pnas.2113751119.Xie L, Cen LP, Li Y, et al. Monocyte-derived SDF1
supports optic nerve regeneration and alters retinal
ganglion cells' response to Pten deletion. Proc Natl Acad
Sci USA. 2022, 119(15): e2113751119. DOI: 10.1073/
pnas.2113751119.
64、u H, Shen B, Han R, et al. CX3CL1-CX3CR1 axis
protects retinal ganglion cells by inhibiting microglia
activation in a distal optic nerve trauma model. Inflamm
Regen. 2024, 44(1): 30. DOI: 10.1186/s41232-024-00343-
4.u H, Shen B, Han R, et al. CX3CL1-CX3CR1 axis
protects retinal ganglion cells by inhibiting microglia
activation in a distal optic nerve trauma model. Inflamm
Regen. 2024, 44(1): 30. DOI: 10.1186/s41232-024-00343-
4.
65、 Zhang ZY, Zuo ZY, Liang Y, et al. Promotion of axon
regeneration and protection on injured retinal ganglion
cells by rCXCL2. Inflamm Regen. 2023, 43(1): 31. DOI:
10.1186/s41232-023-00283-5. Zhang ZY, Zuo ZY, Liang Y, et al. Promotion of axon
regeneration and protection on injured retinal ganglion
cells by rCXCL2. Inflamm Regen. 2023, 43(1): 31. DOI:
10.1186/s41232-023-00283-5.
66、L e i b i n g e r M , M ü l l e r A , A n d r e a d a k i A , e t a l .
Neuroprotective and axon growth-promoting effects
following inflammatory stimulation on mature retinal
ganglion cells in mice depend on ciliary neurotrophic factor
and leukemia inhibitory factor. J Neurosci. 2009, 29(45):
14334-14341. DOI: 10.1523/JNEUROSCI.2770-09.2009.L e i b i n g e r M , M ü l l e r A , A n d r e a d a k i A , e t a l .
Neuroprotective and axon growth-promoting effects
following inflammatory stimulation on mature retinal
ganglion cells in mice depend on ciliary neurotrophic factor
and leukemia inhibitory factor. J Neurosci. 2009, 29(45):
14334-14341. DOI: 10.1523/JNEUROSCI.2770-09.2009.
67、Smith PD, Sun F, Park KK, et al. SOCS3 deletion
promotes optic nerve regeneration in vivo. Neuron. 2009,
64(5): 617-623. DOI: 10.1016/j.neuron.2009.11.021.Smith PD, Sun F, Park KK, et al. SOCS3 deletion
promotes optic nerve regeneration in vivo. Neuron. 2009,
64(5): 617-623. DOI: 10.1016/j.neuron.2009.11.021.
68、Wang D, Luo M, Huang B, et al. Localized co-delivery
of CNTF and FK506 using a thermosensitive hydrogel
for retina ganglion cells protection after traumatic optic
nerve injury. Drug Deliv. 2020, 27(1): 556-564. DOI:
10.1080/10717544.2020.1748759.Wang D, Luo M, Huang B, et al. Localized co-delivery
of CNTF and FK506 using a thermosensitive hydrogel
for retina ganglion cells protection after traumatic optic
nerve injury. Drug Deliv. 2020, 27(1): 556-564. DOI:
10.1080/10717544.2020.1748759.
69、Pernet V, Di Polo A. Synergistic action of brain-derived
neurotrophic factor and lens injury promotes retinal
ganglion cell survival, but leads to optic nerve dystrophy
in vivo. Brain. 2006, 129(Pt 4): 1014-1026. DOI: 10.1093/
brain/awl015.Pernet V, Di Polo A. Synergistic action of brain-derived
neurotrophic factor and lens injury promotes retinal
ganglion cell survival, but leads to optic nerve dystrophy
in vivo. Brain. 2006, 129(Pt 4): 1014-1026. DOI: 10.1093/
brain/awl015.
70、Diamond JS. Inhibitory interneurons in the retina: types,
circuitry, and function. Annu Rev Vis Sci. 2017, 3: 1-24.
DOI: 10.1146/annurev-vision-102016-061345.Diamond JS. Inhibitory interneurons in the retina: types,
circuitry, and function. Annu Rev Vis Sci. 2017, 3: 1-24.
DOI: 10.1146/annurev-vision-102016-061345.
71、Tran NM, Shekhar K, Whitney IE, et al. Single-cell
profiles of retinal ganglion cells differing in resilience to
injury reveal neuroprotective genes. Neuron. 2019, 104(6):
1039-1055.e12. DOI: 10.1016/j.neuron.2019.11.006.Tran NM, Shekhar K, Whitney IE, et al. Single-cell
profiles of retinal ganglion cells differing in resilience to
injury reveal neuroprotective genes. Neuron. 2019, 104(6):
1039-1055.e12. DOI: 10.1016/j.neuron.2019.11.006.
72、Lee S, Chen L, Chen M, et al. An unconventional
glutamatergic circuit in the retina formed by vGluT3
amacrine cells. Neuron, 2014, 84(4): 708-715. DOI:
10.1016/j.neuron.2014.10.021.Lee S, Chen L, Chen M, et al. An unconventional
glutamatergic circuit in the retina formed by vGluT3
amacrine cells. Neuron, 2014, 84(4): 708-715. DOI:
10.1016/j.neuron.2014.10.021.
73、Goldberg JL, Klassen MP, Hua Y, et al. Amacrine-signaled
loss of intrinsic axon growth ability by retinal ganglion
cells. Science. 2002, 296(5574): 1860-1864. DOI:
10.1126/science.1068428.Goldberg JL, Klassen MP, Hua Y, et al. Amacrine-signaled
loss of intrinsic axon growth ability by retinal ganglion
cells. Science. 2002, 296(5574): 1860-1864. DOI:
10.1126/science.1068428.
74、Kunzevitzky NJ, Almeida MV, Duan Y, et al. Foxn4 is
required for retinal ganglion cell distal axon patterning.
Mol Cell Neurosci. 2011, 46(4): 731-741. DOI: 10.1016/
j.mcn.2011.02.004.Kunzevitzky NJ, Almeida MV, Duan Y, et al. Foxn4 is
required for retinal ganglion cell distal axon patterning.
Mol Cell Neurosci. 2011, 46(4): 731-741. DOI: 10.1016/
j.mcn.2011.02.004.
75、Li Y, Andereggen L, Yuki K, et al. Mobile zinc increases
rapidly in the retina after optic nerve injury and regulates
ganglion cell survival and optic nerve regeneration. Proc
Natl Acad Sci USA, 2017, 114(2): E209-E218. DOI:
10.1073/pnas.1616811114.Li Y, Andereggen L, Yuki K, et al. Mobile zinc increases
rapidly in the retina after optic nerve injury and regulates
ganglion cell survival and optic nerve regeneration. Proc
Natl Acad Sci USA, 2017, 114(2): E209-E218. DOI:
10.1073/pnas.1616811114.
76、Trakhtenberg EF, Li Y, Feng Q, et al. Zinc chelation and
Klf9 knockdown cooperatively promote axon regeneration
after optic nerve injury. Exp Neurol. 2018, 300: 22-29.
DOI: 10.1016/j.expneurol.2017.10.025.Trakhtenberg EF, Li Y, Feng Q, et al. Zinc chelation and
Klf9 knockdown cooperatively promote axon regeneration
after optic nerve injury. Exp Neurol. 2018, 300: 22-29.
DOI: 10.1016/j.expneurol.2017.10.025.
77、Zhang Y, Williams PR, Jacobi A, et al. Elevating growth
factor responsiveness and axon regeneration by modulating
presynaptic inputs. Neuron. 2019, 103(1): 39-51.e5. DOI:
10.1016/j.neuron.2019.04.033.Zhang Y, Williams PR, Jacobi A, et al. Elevating growth
factor responsiveness and axon regeneration by modulating
presynaptic inputs. Neuron. 2019, 103(1): 39-51.e5. DOI:
10.1016/j.neuron.2019.04.033.
78、Zhang Q, Xue J, Tang J, et al. Modulating amacrine
cell-derived dopamine signaling promotes optic nerve
regeneration and preserves visual function. Sci Adv. 2024,
10(31): eado0866. DOI: 10.1126/sciadv.ado0866.Zhang Q, Xue J, Tang J, et al. Modulating amacrine
cell-derived dopamine signaling promotes optic nerve
regeneration and preserves visual function. Sci Adv. 2024,
10(31): eado0866. DOI: 10.1126/sciadv.ado0866.
79、Beckers%20A%2C%20Moons%20L.%20Dendritic%20shrinkage%20after%20injury%3A%20%0Aa%20cellular%20killer%20or%20a%20necessity%20for%20axonal%20regeneration%3F.%20%0ANeural%20Regen%20Res.%202019%2C%2014(8)%3A%201313-1316.%20DOI%3A%20%0A10.4103%2F1673-5374.253505.Beckers%20A%2C%20Moons%20L.%20Dendritic%20shrinkage%20after%20injury%3A%20%0Aa%20cellular%20killer%20or%20a%20necessity%20for%20axonal%20regeneration%3F.%20%0ANeural%20Regen%20Res.%202019%2C%2014(8)%3A%201313-1316.%20DOI%3A%20%0A10.4103%2F1673-5374.253505.
80、Beckers A, Van Dyck A, Bollaerts I, et al. An antagonistic
axon-dendrite interplay enables efficient neuronal repair in
the adult zebrafish central nervous system. Mol Neurobiol.
2019, 56(5): 3175-3192. DOI: 10.1007/s12035-018-1292-
5.Beckers A, Van Dyck A, Bollaerts I, et al. An antagonistic
axon-dendrite interplay enables efficient neuronal repair in
the adult zebrafish central nervous system. Mol Neurobiol.
2019, 56(5): 3175-3192. DOI: 10.1007/s12035-018-1292-
5.
81、Cui Q, Yip HK, Zhao RCH, et al. Intraocular elevation of
cyclic AMP potentiates ciliary neurotrophic factor-induced
regeneration of adult rat retinal ganglion cell axons. Mol
Cell Neurosci. 2003, 22(1): 49-61. DOI: 10.1016/s1044-
7431(02)00037-4.Cui Q, Yip HK, Zhao RCH, et al. Intraocular elevation of
cyclic AMP potentiates ciliary neurotrophic factor-induced
regeneration of adult rat retinal ganglion cell axons. Mol
Cell Neurosci. 2003, 22(1): 49-61. DOI: 10.1016/s1044-
7431(02)00037-4.
82、Fournier AE, Takizawa BT, Strittmatter SM. Rho kinase
inhibition enhances axonal regeneration in the injured
CNS. J Neurosci. 2003, 23(4): 1416-1423. DOI: 10.1523/
JNEUROSCI.23-04-01416.2003.Fournier AE, Takizawa BT, Strittmatter SM. Rho kinase
inhibition enhances axonal regeneration in the injured
CNS. J Neurosci. 2003, 23(4): 1416-1423. DOI: 10.1523/
JNEUROSCI.23-04-01416.2003.
83、Drummond ES, Rodger J, Penrose M, et al. Effects of
intravitreal injection of a Rho-GTPase inhibitor (BA-
210), or CNTF combined with an analogue of cAMP, on
the dendritic morphology of regenerating retinal ganglion
cells. Restor Neurol Neurosci. 2014, 32(3): 391-402. DOI:
10.3233/RNN-130360.Drummond ES, Rodger J, Penrose M, et al. Effects of
intravitreal injection of a Rho-GTPase inhibitor (BA-
210), or CNTF combined with an analogue of cAMP, on
the dendritic morphology of regenerating retinal ganglion
cells. Restor Neurol Neurosci. 2014, 32(3): 391-402. DOI:
10.3233/RNN-130360.
84、Francis MM, Freeman MR. Dendrites actively restrain
axon outgrowth and regeneration. Proc Natl Acad
Sci USA. 2016, 113(20): 5465-5466. DOI: 10.1073/
pnas.1605215113.Francis MM, Freeman MR. Dendrites actively restrain
axon outgrowth and regeneration. Proc Natl Acad
Sci USA. 2016, 113(20): 5465-5466. DOI: 10.1073/
pnas.1605215113.
85、Chung SH, Awal MR, Shay J, et al. Novel DLK�independent neuronal regeneration in Caenorhabditis
elegans shares links with activity-dependent ectopic
outgrowth. Proc Natl Acad Sci USA. 2016, 113(20):
E2852-E2860. DOI: 10.1073/pnas.1600564113.Chung SH, Awal MR, Shay J, et al. Novel DLK�independent neuronal regeneration in Caenorhabditis
elegans shares links with activity-dependent ectopic
outgrowth. Proc Natl Acad Sci USA. 2016, 113(20):
E2852-E2860. DOI: 10.1073/pnas.1600564113.
86、Suzuki S, Tachibana M, Kaneko A. Effects of glycine and
GABA on isolated bipolar cells of the mouse retina. J
Physiol. 1990, 421: 645-662. DOI: 10.1113/jphysiol.1990.
sp017967.Suzuki S, Tachibana M, Kaneko A. Effects of glycine and
GABA on isolated bipolar cells of the mouse retina. J
Physiol. 1990, 421: 645-662. DOI: 10.1113/jphysiol.1990.
sp017967.
87、Panter SS, Yum SW, Faden AI. Alteration in extracellular
amino acids after traumatic spinal cord injury. Ann Neurol.
1990, 27(1): 96-99. DOI: 10.1002/ana.410270115.Panter SS, Yum SW, Faden AI. Alteration in extracellular
amino acids after traumatic spinal cord injury. Ann Neurol.
1990, 27(1): 96-99. DOI: 10.1002/ana.410270115.
88、Xu GY, Hughes MG, Ye Z, et al. Concentrations of
glutamate released following spinal cord injury kill
oligodendrocytes in the spinal cord. Exp Neurol. 2004,
187(2): 329-336. DOI: 10.1016/j.expneurol.2004.01.029.Xu GY, Hughes MG, Ye Z, et al. Concentrations of
glutamate released following spinal cord injury kill
oligodendrocytes in the spinal cord. Exp Neurol. 2004,
187(2): 329-336. DOI: 10.1016/j.expneurol.2004.01.029.
89、Romaus-Sanjurjo D, Ledo-García R, Fernández-López B,
et al. GABA promotes survival and axonal regeneration
in identifiable descending neurons after spinal cord injury
in larval lampreys. Cell Death Dis. 2018, 9(6): 663. DOI:
10.1038/s41419-018-0704-9.Romaus-Sanjurjo D, Ledo-García R, Fernández-López B,
et al. GABA promotes survival and axonal regeneration
in identifiable descending neurons after spinal cord injury
in larval lampreys. Cell Death Dis. 2018, 9(6): 663. DOI:
10.1038/s41419-018-0704-9.
90、Goldberg JL, Espinosa JS, Xu Y, et al. Retinal ganglion
cells do not extend axons by default: promotion
by neurotrophic signaling and electrical activity.
Neuron. 2002, 33(5): 689-702. DOI: 10.1016/s0896-
6273(02)00602-5.Goldberg JL, Espinosa JS, Xu Y, et al. Retinal ganglion
cells do not extend axons by default: promotion
by neurotrophic signaling and electrical activity.
Neuron. 2002, 33(5): 689-702. DOI: 10.1016/s0896-
6273(02)00602-5.
91、Li S, Yang C, Zhang L, et al. Promoting axon regeneration
in the adult CNS by modulation of the melanopsin/GPCR
signaling. Proc Natl Acad Sci USA. 2016, 113(7): 1937-
1942. DOI: 10.1073/pnas.1523645113.Li S, Yang C, Zhang L, et al. Promoting axon regeneration
in the adult CNS by modulation of the melanopsin/GPCR
signaling. Proc Natl Acad Sci USA. 2016, 113(7): 1937-
1942. DOI: 10.1073/pnas.1523645113.
92、Lim JH A, Stafford BK, Nguyen PL, et al. Neural activity
promotes long-distance, target-specific regeneration of
adult retinal axons. Nat Neurosci. 2016, 19(8): 1073-1084.
DOI: 10.1038/nn.4340.Lim JH A, Stafford BK, Nguyen PL, et al. Neural activity
promotes long-distance, target-specific regeneration of
adult retinal axons. Nat Neurosci. 2016, 19(8): 1073-1084.
DOI: 10.1038/nn.4340.
93、de Lima S, Koriyama Y, Kurimoto T, et al. Full-length
axon regeneration in the adult mouse optic nerve and
partial recovery of simple visual behaviors. Proc Natl
Acad Sci USA. 2012, 109(23): 9149-9154. DOI: 10.1073/
pnas.1119449109.de Lima S, Koriyama Y, Kurimoto T, et al. Full-length
axon regeneration in the adult mouse optic nerve and
partial recovery of simple visual behaviors. Proc Natl
Acad Sci USA. 2012, 109(23): 9149-9154. DOI: 10.1073/
pnas.1119449109.
94、West AE, Greenberg ME. Neuronal activity-regulated
gene transcription in synapse development and cognitive
function. Cold Spring Harb Perspect Biol. 2011, 3(6):
a005744. DOI: 10.1101/cshperspect.a005744.West AE, Greenberg ME. Neuronal activity-regulated
gene transcription in synapse development and cognitive
function. Cold Spring Harb Perspect Biol. 2011, 3(6):
a005744. DOI: 10.1101/cshperspect.a005744.
95、Yap EL, Greenberg ME. Activity-regulated transcription:
bridging the gap between neural activity and behavior.
Neuron. 2018, 100(2): 330-348. DOI: 10.1016/
j.neuron.2018.10.013.Yap EL, Greenberg ME. Activity-regulated transcription:
bridging the gap between neural activity and behavior.
Neuron. 2018, 100(2): 330-348. DOI: 10.1016/
j.neuron.2018.10.013.
96、Goldstein IM, Ostwald P, Roth S. Nitric oxide: a review of
its role in retinal function and disease. Vision Res. 1996,
36(18): 2979-2994. DOI: 10.1016/0042-6989(96)00017-x.Goldstein IM, Ostwald P, Roth S. Nitric oxide: a review of
its role in retinal function and disease. Vision Res. 1996,
36(18): 2979-2994. DOI: 10.1016/0042-6989(96)00017-x.
97、Aras MA, Aizenman E. Redox regulation of intracellular
zinc: molecular signaling in the life and death of neurons.
Antioxid Redox Signal. 2011, 15(8): 2249-2263. DOI:
10.1089/ars.2010.3607.Aras MA, Aizenman E. Redox regulation of intracellular
zinc: molecular signaling in the life and death of neurons.
Antioxid Redox Signal. 2011, 15(8): 2249-2263. DOI:
10.1089/ars.2010.3607.
98、Spahl DU, Berendji-Grün D, Suschek CV, et al. Regulation
of zinc homeostasis by inducible NO synthase-derived
NO: nuclear metallothionein translocation and intranuclear
Zn2+ release. Proc Natl Acad Sci USA. 2003, 100(24):
13952-13957. DOI: 10.1073/pnas.2335190100.Spahl DU, Berendji-Grün D, Suschek CV, et al. Regulation
of zinc homeostasis by inducible NO synthase-derived
NO: nuclear metallothionein translocation and intranuclear
Zn2+ release. Proc Natl Acad Sci USA. 2003, 100(24):
13952-13957. DOI: 10.1073/pnas.2335190100.